2014 Annual Science Report
 University of Wisconsin
Reporting | SEP 2013 – DEC 2014
University of Wisconsin
Reporting | SEP 2013 – DEC 2014
Project 3E: Sulfur-Cycling Fossil Bacteria From the 1.8 Ga Duck Creek Formation Provide Promising Evidence of Evolution's Null Hypothesis
Project Summary
In the absence of change in the physical-biological environment, evolution of the fundamental aspects of a well-adapted ecosystem — “the form, function and metabolic requirements” of its components — should not occur. Indeed, documentation of evolution in the absence of such changes would show that current understanding of Darwinian evolution is seriously flawed. The mid-Precambrian 2.3 and 1.8 Ga microbial sulfur-cycling assemblages here studied, the first two fossil communities described from quiescent, deep sea, anoxic subsurface mud, are indistinguishable from their modern counterpart. We regard it likely that other essentially identical ancient sub-seafloor microbial biocoenoses will be discovered and think it probable that this initial work will be regarded as having confirmed the linchpin of Darwinian evolution, its logically required null hypothesis.
Project Progress
This work presents five new concepts:
(1) Microbial deep sea sulfur-cycling biotas, “sulfuretums,” are preserved in the geological record (this being only the second such report).
(2) Proliferation of such microbiotas was spurred by the 2.4 -2.2 Ga “Great Oxidation Event” that increased availability of the principal reactants (sulfate and nitrate) which such biotas require.
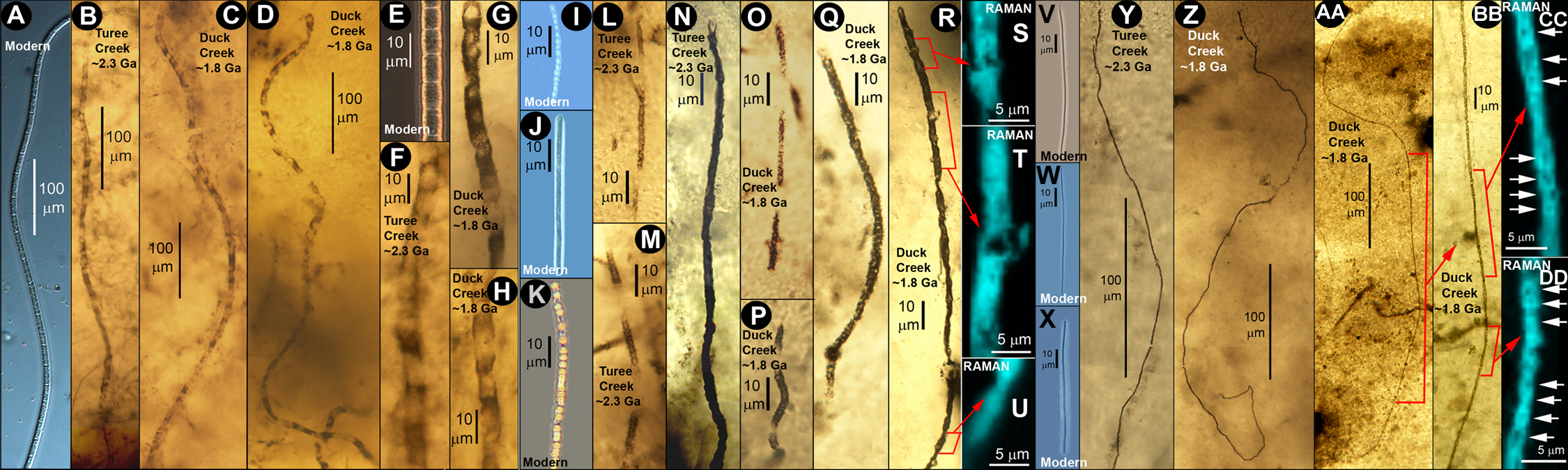
(3) The two mid-Precambrian fossil biotas discussed — separated by a gap of 500 million years — appear to be essentially identical in ecologic setting, fabric, microbial composition, and physiology and are similarly comparable in these respects to living counterpart assemblages, from which they are separated by ~2000 million years.
(4) Because of the stability over geological time of the physically quiescent sub-seafloor anoxic environment inhabited by such biotas, the microbial constituents of such assemblages have not changed fundamentally over the past two billion years (see accompanying figure).
(5) These data are thus consistent with the “negative” null hypothesis required of Darwinian evolution: “if there is no change in the physical-biological environment of a well adapted ecosystem there will be no speciation-based evolution of the form, function or metabolic requirements of its biotic components” — evidence of the validity of Darwin’s theory that can be expected to be provided only by study of an ecosystem fossilized in an environment that has remained essentially unchanged over billions of years.
The logic of our conclusion (Item 5) is a simple syllogism: (1) If Darwinian evolution is a product only of a changing physical-biological environment; (2) and if a changing physical-biological environment does not exist; (3) then such evolution will not occur.
In the absence of such change, evolution of the fundamental aspects of a well-adapted ecosystem — “the form, function and metabolic requirements” of its components — should not occur. Indeed, documentation of evolution in the absence of changes in the physical-biological environment would be an enormously surprising discovery since it would show that current understanding of Darwinian evolution is seriously flawed.
For the first time, this work reports evidence of this “negative” null hypothesis of Darwinian evolution. There are good reasons why it has not previously been documented. This type of evolutionary stasis requires it to be shown in an ecosystem well adapted to an environment that has remained essentially unchanged in its physical and biological characteristics over literally billions of years. This requirement rules out all terrestrial and virtually all marine settings — except for subsurface marine mud in which physical conditions have held steady (due to a lack of wave action, water currents, churning, influx of sediment) and in which its biologically important characteristics (e.g., lack of oxygen, lack of light, lack of a day-night signal, and an influx of required reactants) have remained essentially constant. Similarly, this requirement indicates that relevant evidence should be sought in especially ancient, Precambrian, microbial ecosystems and suggests that such evidence is likely to be provided only by metabolically simple ecosystems in which the components are mutually interdependent and have relatively minimal metabolic demands (for the sulfur-cycling biotas described in the paper, sulfate, sulfide, and nitrate).
Like their modern counterpart, the two microbial assemblages we have studied meet all of these requirements. Although additional data will be needed to establish the sweeping new generalization about Darwinian evolution we propose, we regard it likely that ancient sub-seafloor microbial biocoenoses will be discovered to fill the gap between the mid-Precambrian and the present and that these communities will be fundamentally similar in their form, function and metabolic requirements to those of the Duck Creek and Turee Creek cherts. With the addition of such findings, we think it likely that this initial work will be regarded as having confirmed the null hypothesis of Darwinian evolution, the linchpin required for its universal acceptance.
Publications
-
Schopf, J. W., Kudryavtsev, A. B., Walter, M. R., Van Kranendonk, M. J., Williford, K. H., Kozdon, R., … Flannery, D. T. (2015). Sulfur-cycling fossil bacteria from the 1.8-Ga Duck Creek Formation provide promising evidence of evolution’s null hypothesis. Proc Natl Acad Sci USA, 112(7), 2087–2092. doi:10.1073/pnas.1419241112
-
PROJECT INVESTIGATORS:
-
PROJECT MEMBERS:
James Schopf
Project Investigator
Reinhard Kozdon
Co-Investigator
Anatoliy Kudryavtsev
Co-Investigator
Kenneth Williford
Co-Investigator
Carola Espinoza
Collaborator
David Flannery
Collaborator
Victor Gallardo
Collaborator
John Valley
Collaborator
Martin Van Kranendonk
Collaborator
Malcolm Walter
Collaborator
-
RELATED OBJECTIVES:
Objective 4.1
Earth's early biosphere.
Objective 5.1
Environment-dependent, molecular evolution in microorganisms
Objective 6.1
Effects of environmental changes on microbial ecosystems