2009 Annual Science Report
 Massachusetts Institute of Technology
Reporting | JUL 2008 – AUG 2009
Massachusetts Institute of Technology
Reporting | JUL 2008 – AUG 2009
Genomic Relationships Among Basal Metazoans
Project Summary
The origin of animals, and animal complexity, requires an accurate and precise understanding of both the phylogenetic interrelationship among the earliest evolved animals and the paleoecological milieu within which they evolved. Our work has shown that sponges are indeed the first animals that evolved that still have living descendants, and that sponges are not a natural group. Instead, some sponges are more closely related to more complex animals like humans and jellyfish than they are to other sponges (e.g., bath sponges). This suggests that the origins of animal complexity is rooted in sponge paleobiology, and that the earliest animals were designed to eat bacteria and organic carbon instead of other large eukaryotes like other sponges or plants.
Project Progress
The earliest evidence for animal life comes from the fossil record of 24-isopropylcholestane, a sterane found in Cryogenian deposits, and whose precursors are found in modern demosponges, but not choanoflagellates, calcisponges, hexactinellids or eumetazoans. However, many modern demosponges are also characterized by the presence of siliceous spicules, and there are no convincing demosponge spicules in strata older than the Cambrian. This temporal disparity highlights a problem with our understanding of the Precambrian fossil record – either these supposed demosponge-specific biomarkers were derived from the sterols of some other organism and are simply retained in modern demosponges, or spicules do not primitively characterize crown-group demosponges. Resolving this issue requires resolving the phylogenetic placement of another group of sponges, the hexactinellids, who make a spicule thought to be homologous to the spicules of demosponges, but who also make their first appearance near the Precambrian/Cambrian boundary. Using two independent analytical approaches and data sets – traditional molecular phylogenetic analyses and the presence or absence of specific microRNA genes – we show that demosponges are monophyletic, and that hexactinellids are their sister group (the Silicea). Further, we show that the other two major groups of sponges, homoscleromorphs and calcisponges, are more closely related to eumetaozans than they are to siliceous sponges. Our data suggest that silcecous spicules must have evolved before the last common ancestor of all living siliceans, suggesting the presence of a significant gap in the silicean spicule fossil record. Molecular divergence estimates date the origin of this last common ancestor well within the Cryogenian, consistent with the biomarker record, and strongly suggests that siliceous spicules were present during the Precambrian but are not preserved (Figure 1). On the other hand, the estimate for the age of calcisponges corresponds well with their first appearance in the fossil record (Upper Cambrian, Fig. 1) suggesting that the problem is specific towards a particular biomineral (silica) and a particular clade (Silicea) and not a general perservational failure of all skeletons in the Precambrian.
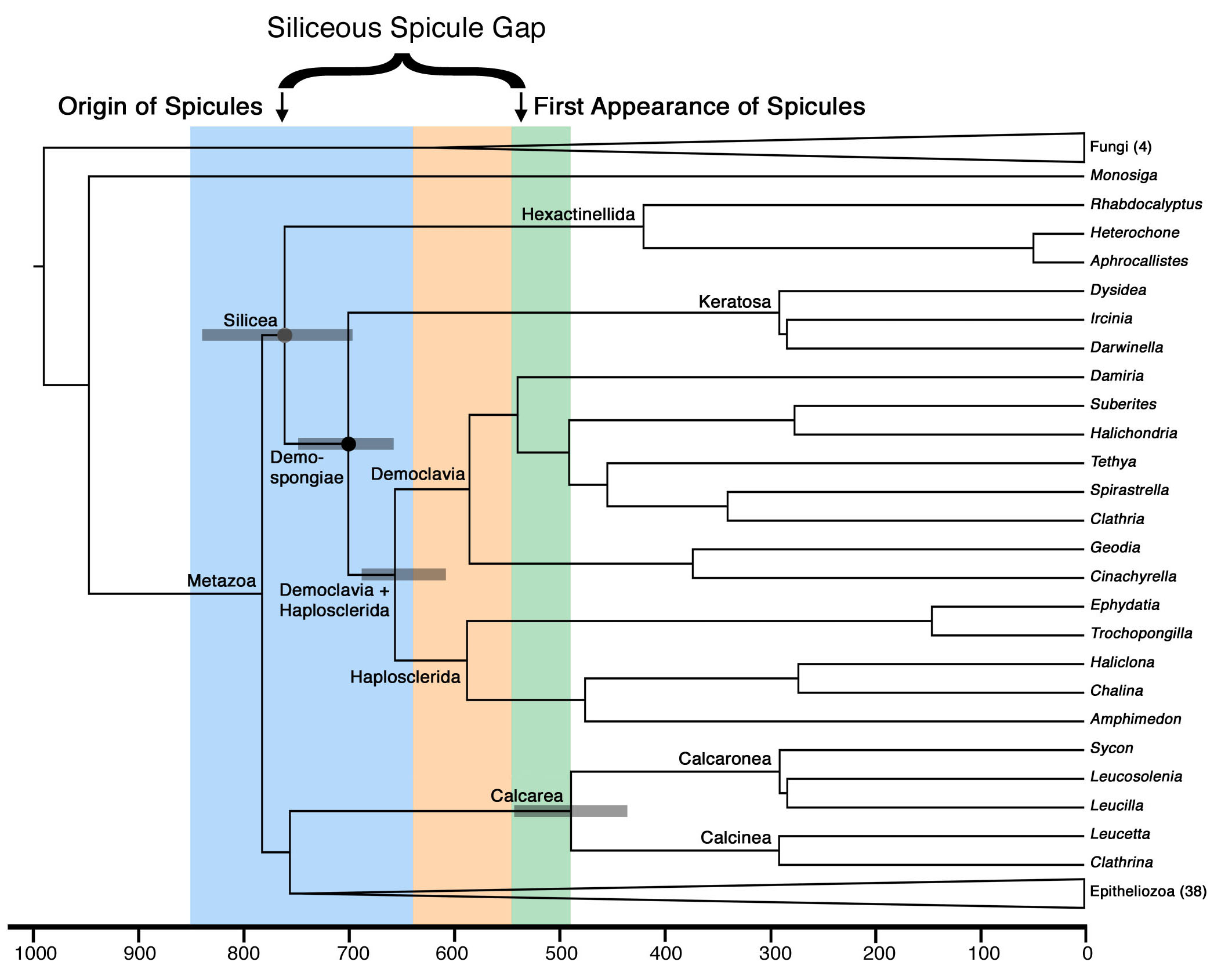
Figure 1. Divergence times estimated under the CIR (rate autocorrelated) relaxed molecular clock model, and the CAT-GTR amino acid substitution model. The phylogeny used to anchor the clock analysis is a pruned version of the phylogeny of Sperling et al. (2009), and includes only higher taxa whose phylogenetic position could be corroborated by the microRNA analysis. According to this analysis, both the origin of metazoans and the diversification of siliceans occurred during the Cryogenian, with further diversification within the spicule-bearing demosponges occurring during the Ediacaran, long before the first appearance of spicule in the fossil record. This disparity highlights a “spicule gap” between their presumed origin in the Cryogenian and their first appearances in the Early Cambrian. Note that our molecular divergence estimates are entirely in agreement with the biomarker record of demosponges (Love et al., 2009) and the fossil record of calcisponges. Ninety-five percent confidence intervals for key nodes are shown in gray boxes.
Publications
-
Sperling, E. A., Robinson, J. M., Pisani, D., & Peterson, K. J. (2010). Where’s the glass? Biomarkers, molecular clocks, and microRNAs suggest a 200-Myr missing Precambrian fossil record of siliceous sponge spicules. Geobiology, 8(1), 24–36. doi:10.1111/j.1472-4669.2009.00225.x
-
PROJECT INVESTIGATORS:
-
PROJECT MEMBERS:
Davide Pisani
Co-Investigator
Scott Nichols
Collaborator
Jeffery Robinson
Doctoral Student
Erik Sperling
Doctoral Student
-
RELATED OBJECTIVES:
Objective 4.2
Production of complex life.
