2015 Annual Science Report
 VPL at University of Washington
Reporting | JAN 2015 – DEC 2015
VPL at University of Washington
Reporting | JAN 2015 – DEC 2015
Astronomical Biosignatures, False Positives for Life, and Implications for Future Space Telescopes
Project Summary
In this task, we identify novel biosignatures and also identify “false positives” for life, which are ways for non-biological processes to mimic proposed biosignatures. Of primary concern are false positives that could mimic easier to detect biosignatures like O2, which we plan to search for with future space-based telescopes. This is a growing area of research that VPL’s past work has motivated, leading to multiple research teams across the planet following our example. Our work continues to be at the forefront of this area of work, as we have identified new non-biological mechanisms for mimicking signs of life. Further, we explained the ways in which these non-biological mechanisms could be identified, and “true positives” from biology confirmed with secondary measurements. Finally, we communicated these lessons to various teams that are studying concepts for future missions that would search for these signs of life. This connection to missions will ensure that our research is incorporated into those missions, so that they will not be “tricked” by these false positives.
Project Progress
This research is divided into two main areas, the development and identification of novel biosignatures, and the in depth study of the robustness of known biosignatures by searching for possible false positives, i.e. planetary processes that may mimic a known biosignature.
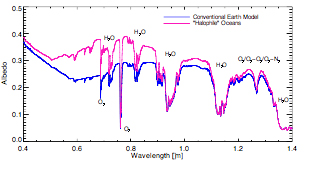
In novel biosignatures, Schwieterman and Meadows collaborated with Cockell of the UK Center for Astrobiology to conduct an interdisciplinary study of the diversity and detectability of non-photosynthetic pigments as biosignatures, especially for halophiles (Schwieterman et al., 2015a). This study included reflectance spectra measurements of a diverse collection of pigmented non-photosynthetic organisms and an analysis of the remote detectability of analogs of these organisms in disk-averaged spectra, with a planetary environment that includes other surfaces, an atmosphere and clouds. These spectra are now available through the existing VPL Biological Pigments Database (http://vplapps.astro.washington.edu/pigments).
In other innovative work on biosignature definition, Krissansen-Totton, Catling and colleague tested the idea that atmospheric chemical disequilibrium can act as a biosignature. They performed the first detailed quantification of thermodynamic disequilibrium in Solar System planetary atmospheres, presenting their work at AbSciCon 2015 (Krissansen Totton et al., 2015 and eventually published as (Krissansen-Totton et al., 2016). The work involved calculating the equilibrium abundances for atmospheric gases using Gibbs free energy minimization, and then calculating the Gibbs energy difference between observed and equilibrium states of an atmosphere to quantify the degree of disequilibrium. They showed that the Earth has ~20 times the thermodynamic chemical disequilibrium of other planets because of the biosphere. They identified that most of the disequilibrium comes from the coexistence of O2, N2 and (liquid) H2O instead of thermodynamically more stable nitric acid. This strong disequilibrium is due to biology: photosynthesis supplies the O2 directly while N2 is produced indirectly through denitrification that uses organic matter.
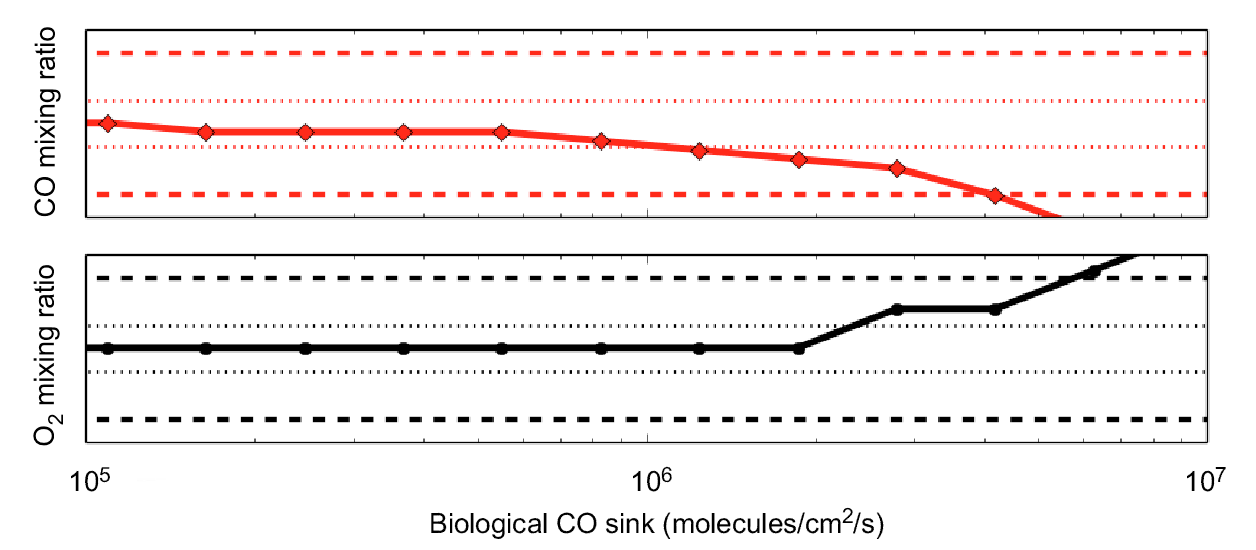
Work is continuing on the topic of biosignatures and disequilibrium with an effort to identify “anti-biosignatures”, where the free energy of abiotic disequilibrium is present but there is no life on a planet’s surface to exploit the free energy. They began by examining the minimum biosphere compatible with carbon monoxide (CO) on Mars, which ordinarily is consumed by a large range of microbes with overall reaction CO + H2O → CO2 + H2. This is done by introducing a biological sink to a 1D Mars photochemical model and finding the maximum possible biological sink consistent with the observed mixing ratios (Fig. 2). This biological sink can then be converted to an approximate biomass. Preliminary results suggest the maximum Martian biomass that could be supported by CO metabolism in the top 1 km of regolith is 0.01 cells/cm3. This biomass is approximately equivalent to less than 100 blue whales across the entire surface of Mars, which is vanishingly small compared to the Earth’s biomass. This work also involved graduate student Steven Sholes, who has been running 1D Mars photochemical models. Future work will examine kinetic measures of disequilibrium rather than thermodynamic ones.
In the area of false positives for previously considered robust biosignatures, this past year, three separate modeling investigations found cases in which O2 and/or O3 could build up abiotically in an exoplanet’s atmosphere. This builds on our work in other tasks (Living Planet, Observer, and Earth Through Time) which has shown us how planetary system components act and interact in complex ways. Gao et al. (2015) used VPL models to show that for a planet orbiting an M dwarf with a high FUV/NUV flux ratio—if the planetary atmosphere is dry and CO2-dominated—then recombination of the products of CO2 photolysis is inhibited. The atmosphere can then build up CO and O2 in 103-104 years, with abundances of abiotic O2 and O3 rivaling that of modern Earth (Gao et al. 2015). In this case, the abiotic production of the O2 is given away by the lack of water in the planetary spectra. However, via the same mechanism of CO2 photolysis, Harman et al. (2015) did a comparison of stellar types, and showed that for planets with liquid water, while abiotic O2 should not accumulate to detectable levels around F and G stars, with K and M stars, the low near-UV flux may allow build-up of O2 if the sinks for O2 are low; meanwhile, O3 could be detectable for a wider range of stars (Harman et al. 2015). Instead of CO2, Luger and Barnes (2015) looked at extreme atmospheric H2O loss during the early high luminosity phase of M dwarfs for planets. For planets that form at the position that will become the 5-Gyr habitable zone, atmospheric evolution is sufficiently severe that the planet can generate large amounts of atmospheric O2 while retaining liquid water (Luger and Barnes, 2015). Thus O2 and O3 detection alone are not robust biosignatures but must be accompanied by knowledge of stellar parameters and a more comprehensive census of atmospheric composition and conditions on terrestrial exoplanets to rule out false positives.
As the direct detection of nitrogen would provide a means to characterize the bulk atmosphere of potentially habitable exoplanets and constrain the likelihood of oxygen production by abiotic processes, Schwieterman et al. (2015) explored a novel way to detect and quantify N2 in planetary atmospheres. Although the N2 molecule is extremely challenging to observe in exoplanet spectra, N2 has a collisional-induced absorption band near 4.2 µm, which is significant in Earth’s spectrum and potentially in those of Earth-like exoplanets with similarly N2-dominated atmospheres. The VPL team quantified the potential magnitude of this spectral signature by producing synthetic transit transmission and radiance spectra using VPL radiative transfer models (Schwieterman et al. 2015b).
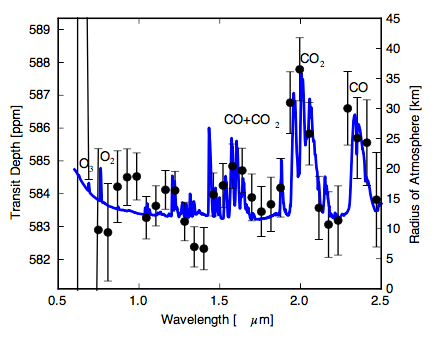
To determine how to best discriminate abiotic accumulation of O2/O3 from photochemical and atmospheric escape, VPL team members Schwieterman, Meadows, Domagal-Goldman, Deming, Arney, Lugar, Harman, Misra and Barnes collaborated on work that produces the first self-consistent transmission spectra of potentially habitable planets with abiotic O2/O3, including estimates of noise using a JWST simulator (Schwieterman et al., 2016). They found that detection of CO (at 2.35, 4.6 um) with CO2 (1.65, 2.0, and 4.3 um) or O4 (1.06, 1.27 um) in transmission spectra would be indicative of abiotic O2 accumulation and could be detected by JWST assuming photon-limited noise. Strong O4 bands seen in reflected light (at 0.345, 0.36, 0.38, 0.445, 0.475, 0.53, 0.57, 0.63, 1.06, and 1.27 um) by future direct-imaging telescopes would indicate an oxygen atmosphere too massive to be biologically produced. This kind of work strengthens our ability to identify true biosignatures by discriminating against the “false positives” when the first spectroscopic studies of rocky habitable zone planets begin.
Finally, we leveraged the knowledge on this task for critical inputs to multiple mission teams, which we anticipate continuing in coming years. This includes the direct participation of VPL team members Meadows and Domagal-Goldman on the Exo-C and Exo-S teams, respectively, and the participation of Domagal-Goldman on the ATLAST Study Team. It also includes presentations by team members Arney, Schwieterman, and Lustig-Jaeger to the ATLAST team. Finally, there were formal publications on telescope architectures and detectors (France et al., 2015; Raucher et al., 2015; Stark et al., 2015) on which we directly participated, or on which our work was acknowledged (e.g., “From Cosmic Birth to Living Earths”, AURA Report, 2015).
Publications
-
Catling, D. C. (2015). Planetary Atmospheres. Treatise on Geophysics, None, 429–472. doi:10.1016/b978-0-444-53802-4.00185-8
-
Gao, P., Hu, R., Robinson, T. D., Li, C., & Yung, Y. L. (2015). STABILITY OF CO 2 ATMOSPHERES ON DESICCATED M DWARF EXOPLANETS. The Astrophysical Journal, 806(2), 249. doi:10.1088/0004-637x/806/2/249
-
Harman, C. E., Schwieterman, E. W., Schottelkotte, J. C., & Kasting, J. F. (2015). ABIOTIC O2LEVELS ON PLANETS AROUND F, G, K, AND M STARS: POSSIBLE FALSE POSITIVES FOR LIFE?. The Astrophysical Journal, 812(2), 137. doi:10.1088/0004-637x/812/2/137
-
Krissansen-Totton, J., Bergsman, D. S., & Catling, D. C. (2016). On Detecting Biospheres from Chemical Thermodynamic Disequilibrium in Planetary Atmospheres. Astrobiology, 16(1), 39–67. doi:10.1089/ast.2015.1327
-
Luger, R., & Barnes, R. (2015). Extreme Water Loss and Abiotic O 2 Buildup on Planets Throughout the Habitable Zones of M Dwarfs. Astrobiology, 15(2), 119–143. doi:10.1089/ast.2014.1231
-
Robinson, T. D., Stapelfeldt, K. R., & Marley, M. S. (2016). Characterizing Rocky and Gaseous Exoplanets with 2 m Class Space-based Coronagraphs. Publications of the Astronomical Society of the Pacific, 128(960), 025003. doi:10.1088/1538-3873/128/960/025003
-
Schwieterman, E. W., Cockell, C. S., & Meadows, V. S. (2015). Nonphotosynthetic Pigments as Potential Biosignatures. Astrobiology, 15(5), 341–361. doi:10.1089/ast.2014.1178
-
Schwieterman, E. W., Meadows, V. S., Domagal-Goldman, S. D., Deming, D., Arney, G. N., Luger, R., … Barnes, R. (2016). IDENTIFYING PLANETARY BIOSIGNATURE IMPOSTORS: SPECTRAL FEATURES OF CO AND O4RESULTING FROM ABIOTIC O2/O3PRODUCTION. The Astrophysical Journal, 819(1), L13. doi:10.3847/2041-8205/819/1/l13
-
Schwieterman, E. W., Robinson, T. D., Meadows, V. S., Misra, A., & Domagal-Goldman, S. (2015). DETECTING AND CONSTRAINING N 2 ABUNDANCES IN PLANETARY ATMOSPHERES USING COLLISIONAL PAIRS. The Astrophysical Journal, 810(1), 57. doi:10.1088/0004-637x/810/1/57
- Stark, C. C., Roberge, A., Mandell, A., Clampin, M., Domagal-Goldman, S. D., McElwain, M. W., & Stapelfeldt, K. R. (2015). Lower limits on aperture size for an ExoEarth-detecting coronagraphic mission. The Astrophysical Journal, 808(2), 149. doi:10.1088/0004-637x/808/2/149
-
PROJECT INVESTIGATORS:
-
PROJECT MEMBERS:
Giada Arney
Co-Investigator
Rory Barnes
Co-Investigator
Robert Blankenship
Co-Investigator
David Catling
Co-Investigator
Mark Claire
Co-Investigator
Chester Harman
Co-Investigator
Tori Hoehler
Co-Investigator
Joshua Krissansen-Totton
Co-Investigator
Rodrigo Luger
Co-Investigator
Amit Misra
Co-Investigator
Niki Parenteau
Co-Investigator
Raymond Pierrehumbert
Co-Investigator
Tyler Robinson
Co-Investigator
Robin Wordsworth
Co-Investigator
Yuk Yung
Co-Investigator
Antigona Segura-Peralta
Collaborator
-
RELATED OBJECTIVES:
Objective 1.1
Formation and evolution of habitable planets.
Objective 1.2
Indirect and direct astronomical observations of extrasolar habitable planets.
Objective 4.1
Earth's early biosphere.
Objective 4.3
Effects of extraterrestrial events upon the biosphere
Objective 5.2
Co-evolution of microbial communities
Objective 5.3
Biochemical adaptation to extreme environments
Objective 7.2
Biosignatures to be sought in nearby planetary systems


