2011 Annual Science Report
 Massachusetts Institute of Technology
Reporting | SEP 2010 – AUG 2011
Massachusetts Institute of Technology
Reporting | SEP 2010 – AUG 2011
Paleontological Investigations of the Advent and Maintenance of Multicellular Life
Project Summary
Understanding the origins and maintenance of complex life requires a two pronged approach: detailed investigations of the ecological and environmental context of the advent of complex life, as revealed by the fossil record, and exploration of the molecular underpinnings of how life becomes complex, how it is maintained, and how it is lost. The appearance of complex life that begins in the Ediacaran period around 580 million years ago and truly blossoms in the Cambrian Period about 530 million years ago reveals that ecological interactions – specifically predation – was a necessary component, and was depended upon the continuing oxygenation of the world’s oceans. Molecular investigations have revealed the genes and gene interactions that appear to be necessary for the advent of complex life, and what needs to be lost in order for complex life to become secondarily simplified. Together the fossil record and the molecular record indicate that evolving complex life involves both new genes and new ecologies within the context of permissive environmental circumstances.
Project Progress
Paleontological Investigations of the Advent and Maintenance of Multicellular Life
The advent and early evolution of multicellular life involves interactions between environmental potential, the possibilities provided by genetic and developmental innovations, and the construction of ecological opportunities. Our NAI group has focused on investigations of each of the three elements of this macroevolutionary triad, and on the interactions between them.
Graduate Student Sarah Tweedt and Doug Erwin compiled a database of the first occurrences of all metazoan phyla, classes, orders and equivalent stem clades through the Phanerozoic, but with particular emphasis on the Ediacaran-Ordovician. Although there have been incredible fossil discoveries over the past couple of decades, new phylogenetic relationships established, and a wholesale revision of the timescale, no such compilation had been made since the late 1980s. The results further emphasize the rapidity and magnitude of the Cambrian Explosion. Significantly, the many stem-clades that have been recognized do not form intermediates between previously recognized clades but further enhance morphologic disparity. This compilation has served as the basis for a number of other projects.
This database of occurrences, in combination with other fossil data was employed in a study of the role of ecosystem engineering during the Cambrian explosion. Ecosystem engineering is the modification of resources by the physical and chemical effects of organism activities, including modification of the sediment by burrowing, construction of reefs, and other activities. Erwin and Tweedt analyzed the pattern of ecosystem engineering from the Ediacaran through the Cambrian and showed a major increase associated with the early Cambrian. Much of this increase was associated with processes that are expected to have significant ecological spillover, and thus generate positive feedback for further increases in diversity.
Post-doctoral fellow Marc Laflamme expanded this database to include the occurrences of Ediacara macrofossils and produced a new classification of these enigmatic fossils. His revised classification scheme highlights a new framework for Ediacaran phylogeny and classification that emphasizes branching or segment architecture, body symmetry, associated trace fossils, and growth parameters, while restricting direct comparisons with modern taxa unless they share undoubted synapomorphies. Care was taken to ensure that groupings were not representative of taphonomic, biogeographic, paleoecological, or taxonomic biases, which have overprinted previous attempts to classify these organisms. This resulted in the establishment of six clades and four likely clades that can account for the majority of diversity of Ediacaran form, and allows for the classification of 64 species (out of 89 valid species) into distinct higher-order clades.
These clades were then used to tackle the question of an implied Ediacaran extinction at the onset of the Cambrian explosion of metazoans (especially bilaterians). Clade spatial distributions were investigated by plotting their modern distributions onto paleocontinental reconstructions from the Ediacaran using Christopher Scotese’s Paleomap software. Several clades were demonstrated to have spanned the entire Ediacaran, disappearing from the fossil record prior to the Cambrian explosion.
In a Research Article currently in press at Science, we integrated this fossil data with new molecular clock data from Principle Investigator Kevin Peterson and his colleagues to generate a new synthesis of the pattern and causes of the Cambrian Explosion. In this paper we show that the major animal clades diverged many tens of millions of years before their first appearance in the fossil record, demonstrating a macroevolutionary lag between the establishment of their developmental toolkits during the Cryogenian (850-635 Ma), and the later ecological success of metazoans during the Ediacaran (635-541 Ma) and Cambrian (541-488 Ma) periods. We argue that this diversification involved new forms of developmental regulation, as well as innovations in networks of ecological interaction within the context of permissive environmental circumstances.
Team member Kevin Peterson and colleagues focused primarily on the relationship between morphological complexity/simplicity and the underlying developmental tool kit, specifically microRNAs. microRNAs (miRNAs) are small, ~22 nt non-coding RNA genes that impact the translation of messenger RNAs, and thus are genetic regulators that function post-transcriptionally in the cytoplasm. We have shown (Heimberg et al. 2010) that there is a significant correlation between the advent of morphological complexity, as measured by the advent of novel cell types, and a dramatic increase in the number of discrete miRNAs. Specifically, we showed that at the base of vertebrates, which constitutes one of the greatest increases to morphological complexity in the history of animal evolution, there is a dramatic increase in the number of miRNA families. Importantly, these vertebrate-specific miRNAs are expressed at very high levels in vertebrate-specific tissues – for example the highest expressed miRNA in the vertebrate liver, an organ not found in any invertebrate, is the vertebrate-specific miRNA miR-122. On the other hand, instances of secondary morphological simplification are associated with dramatic reductions to the animal’s miRNA repertoire. In work published in Nature earlier this year (Philippe et al. 2011), we showed that animals whose relatively simple morphology (few discrete cell types and absence of discreet organs) have suggested to most workers a deep position on the animal tree of life are instead secondarily simplified deuterostomes, the great clade of animals that includes echinoderms (sea urchins and starfish) and vertebrates including humans. Specifically we were able to show based on considerations of both miRNAs and phylogenomics that acoel flatworms, along with their close relatives nemertodermatids and Xenoturbella, are closely related to the ambulacrarian deuterostomes (echinoderms and hemichordates). Elsewhere on the animal tree, where there is little increase to morphological complexity but also no secondary simplification, miRNAs are added over time but at a greatly reduced rate as compared to the base of vertebrates, for example (e.g., Campbell et al. 2011; Campo-Paysaa et al. 2011; Pisani et al., 2011; Sperling et al. 2011). Thus, because of the affect miRNAs have on gene expression, and the role they might play in the evolution of novel cell types, miRNAs might be instrumental in the evolution of complex morphologies, at least within animals. This work is summarized and placed into paleobiological context in the Erwin et al. paper currently in press at Science.
With graduate student Jeffery Robinson, team member Peterson has also focused on trying to better understand the role miRNAs play in the advent of multicellularity by exploring the role miRNAs play in sponge development and cellular homeostatic maintenance. Specifically we are addressing whether miRNAS in sponges show any sort of developmental regulation (more complex animals show that most miRNAs are expressed relatively late as they are responsible for maintaining cellular differentiation), and whether sponges show cell type enrichment for specific genes (like vertebrates do for example, with liver cells copiously expressing miR-122). To explore the temporal regulation of miRNAs through developmental time, the freshwater demosponge Ephydatia mulleri was collected as gemmules (resistant capsules of undifferentiated stem cells known as thesocytes), and allowed to activate and differentiate into small juvenile sponges under laboratory conditions. 454 small RNA libraries were then made from gemmule, newly differentiated juvenile, and adult sponge tissue collected at the same site as the gemmules. All material was identified to species level morphologically by spicule preparation, and molecular identification using 18S, CO1, and ITS2 sequences PCR’d from collected material and compared to database accessions of type specimens. 454 libraries were sequenced, and showed that sponge mir-2016 is expressed predominantly in gemmule and newly differentiated juvenile tissue, while mir-2019 is expressed in differentiated adult tissue. Thus, like in complex bilaterian animals, sponges too show differential regulation of miRNAs through development. To explore the second issue, that of differential spatial regulation of miRNAs, the marine demosponge Suberites sp. was obtained from biological suppliers and maintained in saltwater aquaria. Ficoll density gradient centrifugation and flow cytometry protocols were developed and used to isolate specific cell types including archaeocytes and choanocytes, and the RNA derived from these cell-enriched populations are being prepared for Solexa sequencing. Similar to our temporal regulation results, we expect to see that different miRNAs will show differential spatial regualation as well, consistent with the idea that cell-type complexity relies on a differential miRNA expression profile.
Sarah Tweedt, Doug Erwin, and Marc Laflamme are presently collaborating on the construction of a theoretical morphospace of the Ediacara biota. Many Ediacarans consist of self-repeating (fractal) modules that are unique to these organisms. In the absence of modern examples, biological aspects of these modular constructions must be studied through computer modeling. Modeling allows for direct manipulation of growth parameters and organismal construction. Through directed manipulation of the available morphological parameters and growth constraints as established from the fossil record, it is possible to reconstruct a theoretical morphospace of known, likely, unknown, and even impossible constructions, which will significantly aid in future fossil reconstruction, provide novel search images for paleontologists, and ultimately isolate which environmental and behavioral constraints (i.e. nutrient acquisition, substrate stability, etc.) were most important in constraining evolutionary novelty. Biological efficiency can be tested via surface-area to volume measurements, which represent a proxy for absorptive efficiency necessary in Ediacaran feeding. Finally, a data matrix of morphological characters that highlight inferred behavioural mechanisms (predation, filter feeding, etc.) that affect ecosystem construction will be investigated in order to study the paleoecological transition in ecosystem construction at the onset of complex animals.
Team member Andrew Knoll also focused on the evolution of complex multicellularity, with efforts in three directions. Much of the Knoll lab’s work involved geochemical attempts to constrain the Neoproterozoic-Cambrian evolution of oxygen and redox profiles in the oceans. Mo isotopes support a growing body of data suggesting that anoxic water masses existed in shallow (no more than a few tens of meters) oceans as recently as 750 million years ago (Dahl et al., in press). Moreover, although several lines of evidence indicate that shallow oceans became more generally oxic during the Ediacaran Period, widespread subsurface dysoxia persisted in or returned to the world’s oceans during the Cambrian Period (Gill et al., 2011), again supporting other lines of evidence that suggest intermediate and not modern, levels of oxygen in the Early Paleozoic oceans where animals diversified. The Mo isotope research reflects collaboration with another NAI team (Anbar, ASU). In other research, the Knoll group completed a paper documenting secular variations in Si isotopic abundance through Precambrian time, providing a new avenue for investigating both the redox structure of ancient oceans and the changing proportional contributions of hydrothermal activity and continental weathering in governing sea water chemistry (Chakrabrarti et al., submitted). The group also completed a first round of experiments designed to provide physiological tests of geology-based hypotheses for major changes through time in Earth’s biota – in this case we tested hypotheses which posit that seawater composition influenced the evolution of the phytoplankton on which complex marine organisms ultimately depend (Ratti et al., 2011). One additional paper examined evidence for environmental controls on the evolution of skeletonized marine animals through time, presenting a new hypothesis for the episodic expansion and decline of metazoan reefs through time (Knoll and Fischer, 2011).
In a second line of research, the Knoll group has made substantial progress in understanding at a mechanistic level the preservation of early animal skeletons by phosphatization (Creveling et al., 2011) and the preservation of non-mineralized animals as carbonaceous compressions (Burgess Shale-type preservation, Gill et al., 2011). These preservational mechanisms together account for much of our insights into early animal evolution; this research continues and will be a primary focus of research in the coming year.
Knoll’s third line of research involved collaboration with molecular biologists in attempt integrate fossils and phylogeneis to provide meolcular clock estimates of major events in eukaryotic evolution. In one paper, Parfrey et al. (2011) indicated that the last common ancestor of extant eukaryotes most likely existed 1800-1600 million years ago. In a detailed phylogeteic analysis of red alage – which evolved complex mutlicelluairty independently of plants and animals, Wang et al. showed, in complementary fashion that red algae diverged from greens more than a billion years ago, that complex red algae (Florideophytes) evolved 700-600 million years ago, and that a major radiation within the Florideophytes substantially increased red algal diversity during the mid-Paleozoic Era (Yang et al., submitted). Finally, Knoll published a review comparing the evolution of complex multicellularity in different clades (Knoll, 2011)
Team members Erwin and Knoll both completed book projects. Erwin continued work on his book on the Cambrian Explosion with Jim Valentine, which should be published in 2012. Knoll (along with Donald Canfield and Kurt Konhauser) wrapped up an edited introduction to geobiology, also to be published in 2012. Team member Peterson was put under contract by John Wiley and Sons for a multi-author work on Molecular Paleobiology.
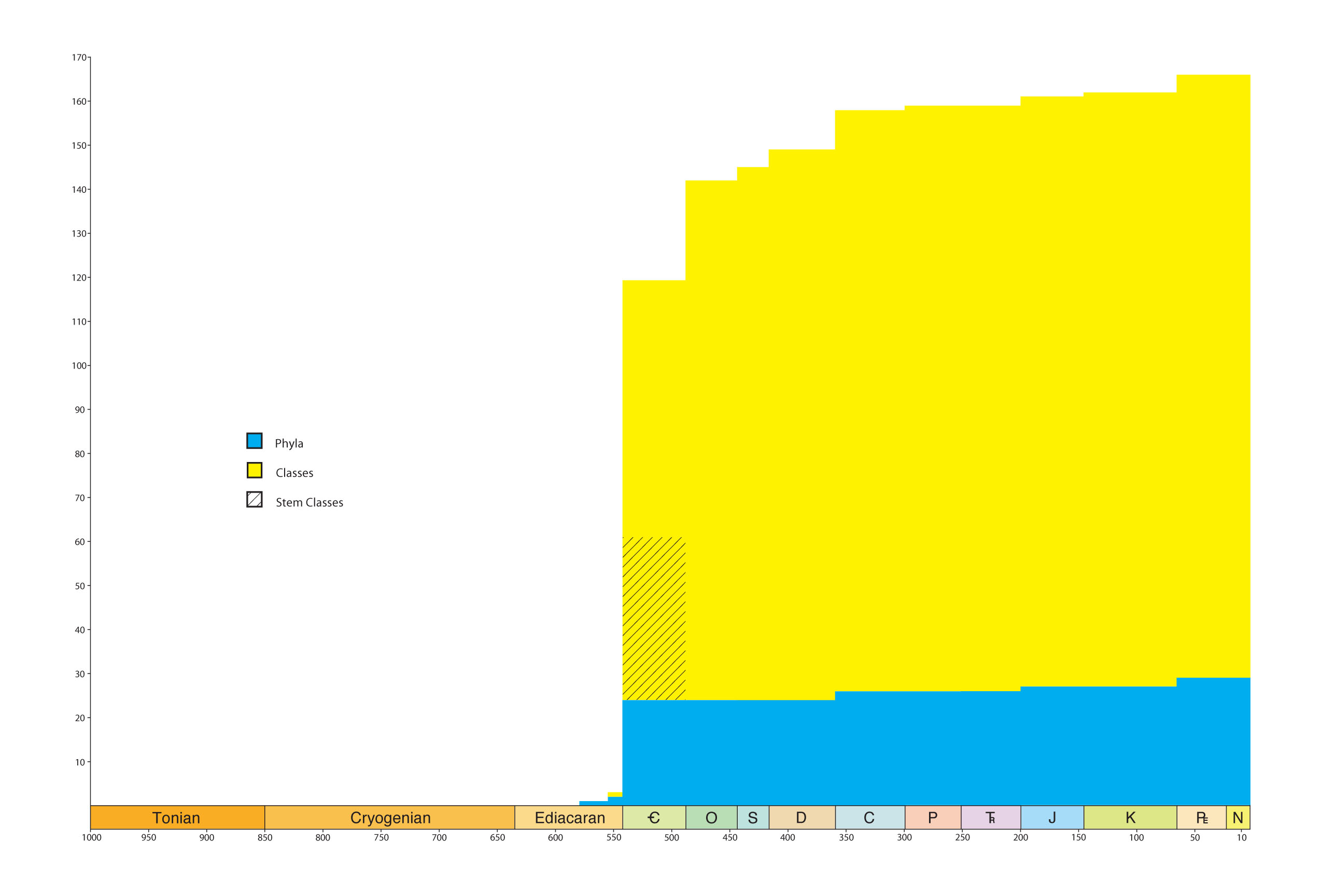
Diversity of phyla (blue), classes (yellow) and stem clades (hatched) through the Phanerozoic, demonstrating the overwhelming impact of the Cambrian explosion on the diversity and disparity of metazoan life.
Publications
-
Campbell, L. I., Rota-Stabelli, O., Edgecombe, G. D., Marchioro, T., Longhorn, S. J., Telford, M. J., … Pisani, D. (2011). MicroRNAs and phylogenomics resolve the relationships of Tardigrada and suggest that velvet worms are the sister group of Arthropoda. Proceedings of the National Academy of Sciences, 108(38), 15920–15924. doi:10.1073/pnas.1105499108
-
Campo-Paysaa, F., Sémon, M., Cameron, R. A., Peterson, K. J., & Schubert, M. (2011). microRNA complements in deuterostomes: origin and evolution of microRNAs. Evolution & Development, 13(1), 15–27. doi:10.1111/j.1525-142×.2010.00452.x
-
Chakrabarti, R., Knoll, A. H., Jacobsen, S. B., & Fischer, W. W. (2012). Si isotope variability in Proterozoic cherts. Geochimica et Cosmochimica Acta, 91, 187–201. doi:10.1016/j.gca.2012.05.025
-
Dahl, T. W., Canfield, D. E., Rosing, M. T., Frei, R. E., Gordon, G. W., Knoll, A. H., & Anbar, A. D. (2011). Molybdenum evidence for expansive sulfidic water masses in ~750Ma oceans. Earth and Planetary Science Letters, 311(3-4), 264–274. doi:10.1016/j.epsl.2011.09.016
-
Erwin, D. H. (2011). Evolutionary uniformitarianism. Developmental Biology, 357(1), 27–34. doi:10.1016/j.ydbio.2011.01.020
-
Erwin, D. H. (2011). Novelties That Change Carrying Capacity. J. Exp. Zool. (Mol. Dev. Evol.), 318(6), 460–465. doi:10.1002/jez.b.21429
-
Erwin, D. H., & Tweedt, S. (2011). Ecological drivers of the Ediacaran-Cambrian diversification of Metazoa. Evol Ecol, 26(2), 417–433. doi:10.1007/s10682-011-9505-7
-
Erwin, D. H., Laflamme, M., Tweedt, S. M., Sperling, E. A., Pisani, D., & Peterson, K. J. (2011). The Cambrian Conundrum: Early Divergence and Later Ecological Success in the Early History of Animals. Science, 334(6059), 1091–1097. doi:10.1126/science.1206375
-
Gill, B. C., Lyons, T. W., Young, S. A., Kump, L. R., Knoll, A. H., & Saltzman, M. R. (2011). Geochemical evidence for widespread euxinia in the Later Cambrian ocean. Nature, 469(7328), 80–83. doi:10.1038/nature09700
-
Heimberg, A. M., Cowper-Sal{middle Dot}lari, R., Semon, M., Donoghue, P. C. J., & Peterson, K. J. (2010). microRNAs reveal the interrelationships of hagfish, lampreys, and gnathostomes and the nature of the ancestral vertebrate. Proceedings of the National Academy of Sciences, 107(45), 19379–19383. doi:10.1073/pnas.1010350107
-
Knoll, A. H. (2011). The Multiple Origins of Complex Multicellularity. Annual Review of Earth and Planetary Sciences, 39(1), 217–239. doi:10.1146/annurev.earth.031208.100209
-
Laflamme, M., Flude, L. I., & Narbonne, G. M. (2012). Ecological Tiering and the Evolution of a Stem: The Oldest Stemmed Frond from the Ediacaran of Newfoundland, Canada. Journal of Paleontology, 86(2), 193–200. doi:10.1666/11-044.1
-
Laflamme, M., Schiffbauer, J. D., Narbonne, G. M., & Briggs, D. E. G. (2010). Microbial biofilms and the preservation of the Ediacara biota. Lethaia, 44(2), 203–213. doi:10.1111/j.1502-3931.2010.00235.x
-
Parfrey, L. W., Lahr, D. J. G., Knoll, A. H., & Katz, L. A. (2011). Estimating the timing of early eukaryotic diversification with multigene molecular clocks. Proceedings of the National Academy of Sciences, 108(33), 13624–13629. doi:10.1073/pnas.1110633108
-
Philippe, H., Brinkmann, H., Copley, R. R., Moroz, L. L., Nakano, H., Poustka, A. J., … Telford, M. J. (2011). Acoelomorph flatworms are deuterostomes related to Xenoturbella. Nature, 470(7333), 255–258. doi:10.1038/nature09676
-
Pisani, D., Feuda, R., Peterson, K. J., & Smith, A. B. (2012). Resolving phylogenetic signal from noise when divergence is rapid: A new look at the old problem of echinoderm class relationships. Molecular Phylogenetics and Evolution, 62(1), 27–34. doi:10.1016/j.ympev.2011.08.028
-
Ratti, S., Knoll, A. H., & Giordano, M. (2011). Did sulfate availability facilitate the evolutionary expansion of chlorophyll a+c phytoplankton in the oceans?. Geobiology, 9(4), 301–312. doi:10.1111/j.1472-4669.2011.00284.x
-
Sperling, E. A., Peterson, K. J., & Laflamme, M. (2010). Rangeomorphs, Thectardis (Porifera?) and dissolved organic carbon in the Ediacaran oceans. Geobiology, 9(1), 24–33. doi:10.1111/j.1472-4669.2010.00259.x
-
Sperling, E. A., Pisani, D., & Peterson, K. J. (2011). Molecular paleobiological insights into the origin of the Brachiopoda. Evolution & Development, 13(3), 290–303. doi:10.1111/j.1525-142×.2011.00480.x
- Bush, A.M., Bambach, R.K. & Erwin, D.H. (2011). Ecospace utilization during the Ediacaran radiation and the Cambrian Eco-plosion. In: LaFlamme, M., Schiffbauer, J. & Dornbos, S. (Eds.). Quantifying the Evolution of Early Life. Vol. Topics in Geobiology. Springer.
- Creveling, J., Johnston, D.T. & Knoll, A.H. (2011). Geochemical controls on phosphatization taphonomy in the Middle Cambrian. Geological Society of America, 43(5): 53.
- Darroch, S., Laflamme, M., Schiffbauer, J.D. & Briggs, D.E.G. (Accepted). Experimental Formation of a Microbial Death Mask. Palaios.
- Dornbos, S.Q., Clapham, M.E., Fraiser, M.L. & Laflamme, M. (In Press). Lessons from the Fossil Record: The Ediacaran Radiation, the Cambrian Radiation, and the End-Permian Mass Extinction [Book Chapter]. Marine Biodiversity Futures and Ecosystem Functioning frameworks, methodologies and integration.
- Erwin, D.H. & Valentine, J.W. (In Presss). The Cambrian Explosion: The Construction of Animal Biodiversity. Ben Roberts.
- Erwin, D.H. (2011). A Paleontologist looks at history. Cliodynamics, 2: 27-39.
- Gill, B., Knoll, A.H. & Johnston, D.T. (2011). Investigating geochemistry of the Wheeler Formation of Utah, USA: Insights into the redox environment of Burgess Shale-type fossil preservation. Geological Society of America, 43(5): 53.
- Knoll, A.H. & Fischer, W.W. (2011). Knoll, A.H. and W.W. Fischer. In: J.P. Knoll, A.H.a.W.W.F. (Eds.). Ocean Acidification. Oxford University Press.
- Knoll, A.H. & Konhauser, D.E.C.a.K. (2011, In Press). Essentials of Geobiology. In: Knoll, A.H. & Konhauser, D.E.C.a.K. (Eds.). Cambridge MA: Wiley-Blackwell.
- Laflamme, M. & Casey, M.M. Morphometrics in the study of Ediacaran fossil shapes. In: Laflamme, M., Schiffbauer, J.D., Dornbos & Springer’s, S.Q. (Eds.). Quantifying the Evolution of Early Life: Numerical and Technological Approaches to the Evaluation of Fossils and Ancient Ecosystems. Vol. 36.
- Laflamme, M., Schiffbauer, J.D. & Narbonne, G.M. (In Press). Deep-Water Microbially Induced Sedimentary Structures (MISS) in Deep Time: The Ediacaran Fossil Ivesheadia in Microbial Mats in Sandy Deposits (Archean Era to Today). In: Noffke and Henry Chafetz, N.K. (Eds.). SEPM Special Publication.
- Pruss, S., Clemente, H. & Laflamme, M. (Accepted). Early (Series 2) Cambrian archaeocyathid reefs as a locus for skeletal carbonate production: New insights from the Forteau Formation, southern Labrador. Lethaia.
- Yang, E.C., Boo, S.M., Bhattacharya, D., Saunders, G.W., Knoll, A.H., Fredericq, S. & Yoon, H.S. (2011, Submitted). Divergence time estimates and evolution of major lineages in the florideophyte red algae. PLoS One.
-
PROJECT INVESTIGATORS:
-
PROJECT MEMBERS:
Davide Pisani
Collaborator
Marc LaFlamme
Postdoc
Erik Sperling
Postdoc
Sarah Tweedt
Doctoral Student
Jeffery Robinson
Graduate Student
Jonathan Wilson
Graduate Student
-
RELATED OBJECTIVES:
Objective 4.2
Production of complex life.


