
Nitrogen is a major nutrient for all terrestrial life. Life’s distribution, abundance and evolution is thus dependent on the bio-availability of this essential element, but conversely life’s evolution has modified the distribution, speciation and abundance of nitrogen. As a result, the nitrogen cycle has changed markedly through time.
In the Hadean and Eoarchean, abiotic fixation would have allowed a very small flux of bio-available nitrogen from the atmosphere, severely limiting the size and diversity of the primordial biosphere. However, efficient biological fixation using Mo-nitrogenase was established by the Mesoarchean, leading to large biomass deposition in marine settings and (possibly) depletion of nitrogen from the atmosphere as a result of enhanced organic burial. During the late Neoarchean in conjunction with the first whiffs of biogenic oxygen to the atmosphere, aerobic nitrogen cycling was temporarily initiated, marked by positive fractionations of nitrogen isotopes indicating active denitrification sending nitrogen gas back to the atmosphere.
The Paleoproterozoic “Great Oxidation Event” and subsequent “oxygen overshoot” during the Lomagundi carbon isotopic excursion were marked by widespread aerobic cycling with vigorous denitrification. Towards the end of the Paleoproterozoic and through the Mesoproterozoic, expanded euxinic (sulfidic) conditions in the oceans during the “boring billion” years of environmental stasis caused spatial perturbations in the nitrogen cycle, with aerobic cycling near-shore but anaerobic fixation-dominated cycling offshore. As sulfide strips dissolved copper from seawater, thus removing the key cofactor for the critical enzyme in the last step of denitrification, a nitrous oxide greenhouse may have been operative at this time. It is not clear when the nitrogen cycling attained its modern fully aerobic aspect, but it may have been as late as the Devonian when the deep oceans may have finally become permanently oxidized.
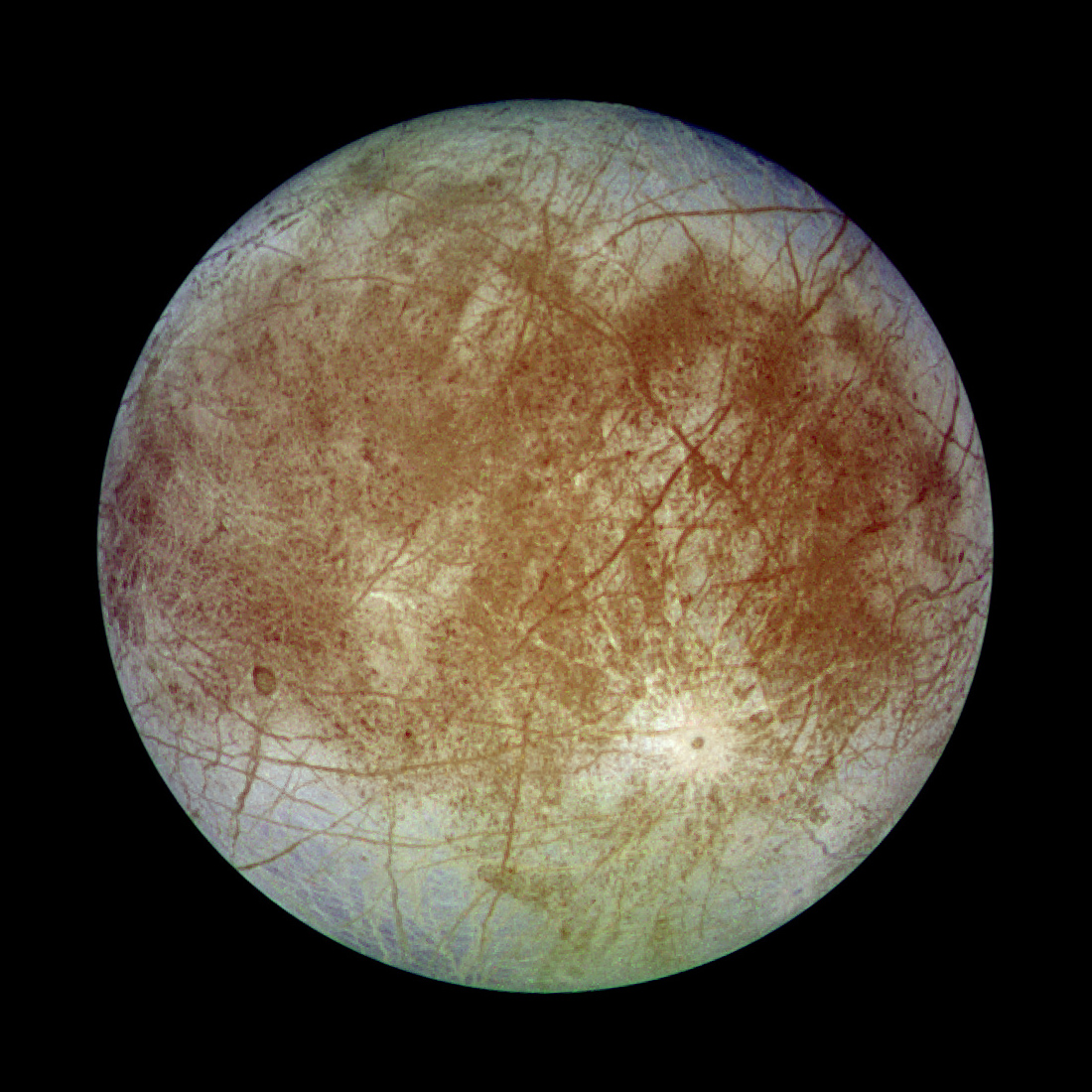 Getting Under Europa’s Skin
Getting Under Europa’s Skin Tracing Formation and Evolution of Outer Solar System Bodies Through Stable Isotopes and Noble Gas Abundances
Tracing Formation and Evolution of Outer Solar System Bodies Through Stable Isotopes and Noble Gas Abundances Photosynthesis, a Planetary Revolution
Photosynthesis, a Planetary Revolution Xenon: King of the Gases
Xenon: King of the Gases