2015 Annual Science Report
 University of California, Riverside
Reporting | JAN 2015 – DEC 2015
University of California, Riverside
Reporting | JAN 2015 – DEC 2015
Executive Summary
A single question drives the research of the Alternative Earths Team of the NASA Astrobiology Institute (NAI): How has Earth remained persistently inhabited through most of its dynamic history, and how do those varying states of inhabitation manifest in the atmosphere? Simply put, we are unraveling the evolving redox state of Earth’s early atmosphere as a guide for exoplanet exploration. Atmospheric redox and the abundance of associated gases are fingerprints of the complex interplay of processes on and within a host planet that point both to the presence and possibility of life. Redox-sensitive greenhouse gases, for example, can expand the habitable zone well beyond what is predicted from the size of a planet’s star and its distance from that energy source alone. Conversely, the absence of obvious biosignature gases such as oxygen does not necessarily mean a planet is sterile: cyanobacteria were producing oxygen on Earth long before it accumulated to remotely detectable concentrations in the atmosphere.
During Year 1, the Alternative Earths team was active with productive collaboration among co-investigators at UC Riverside (UCR), Yale, Georgia Tech (GT), Arizona State University (ASU), Oregon Health and Science University (OHSU), and the J. Craig Venter Institute (JCVI); with our collaborators at home and abroad (Penn State, Rice University, Aarhus University, University of Southern Denmark, and Rhodes University); and through emerging synergism with other NAI teams. Anchoring our first wave of research results are the fruits of extensive lab- and fieldwork as well as first steps toward our ultimate goal of modeling early Earth atmospheres, beginning with the mid-Proterozoic. Indeed, our latest modeling of biosignature gases in the mid-Proterozoic atmosphere is revealing intriguing implications for climate stability and ‘false negatives’ in remote life detection—despite the earliest emergence of complex life in the oceans below.
To organize our comprehensive deconstruction of the geologic record, from the earliest biological production of oxygen to its permanent accumulation in large amounts almost three billion years later, we originally selected three critical time intervals centered on a compelling question or controversy:
* Alternative Earth 1 – In the mid-late Archean (3.2 to 2.4 billion years ago) we are searching the rock record for the earliest atmospheric and oceanic traces of oxygenic photosynthesis and the reasons behind their timing.
* Alternative Earth 2 – In the mid-Paleoproterozoic (2.2 to 2.0 billion years ago) we are investigating whether Earth’s surface experienced a unidirectional oxygen rise or instead rose to near-modern levels, then crashed dramatically.
* Alternative Earth 3 – In the mid-Proterozoic (1.8 to 0.8 billion years ago), we are exploring the interplay among oxygen, the rise and increasing complexity of eukaryotes, and conditions that set the stage for the eventual rise of metazoans.
Over the course of Year 1, we also nurtured our natural synergism with the Origins of Complexity NAI Team. The result is a new time interval of primary interest:
* Alternative Earth 4 – In the late Proterozoic (0.8 to 0.5 billion years ago), our central goal is to work with the Origins of Complexity NAI Team to better understand how the rise of complex life shaped planetary-scale biosignatures.
Research Highlights
Compelling progress in all four of our Alternative Earth research categories is summarized in our 2015 Project Reports. Here we feature one redox proxy in particular, due to its great utility in addressing atmospheric oxygen levels spanning all four Alternative Earth time intervals. The chromium (Cr) isotope proxy tracks periods when atmospheric oxygen was sufficiently high to support coupled oxidative cycles for manganese (Mn) and Cr in ancient soils. The critical benefit is that this tracer is sensitive to pO2 levels a few orders of magnitude higher than those required to turn off the preservation of non-mass-dependent sulfur isotope fractionation. As such, Cr isotopes can fingerprint levels of intermediate oxygenation that might manifest during transient ‘whiffs of oxygen’ in the otherwise oxygen-free Archean, for instance. Our results also show that Cr isotopes are particularly useful for tracing the fabric of oxygenation during the rise of eukaryotes following the Great Oxidation Event.
This work comprises more than 300 new Cr isotope measurements from 20 different geologic formations and shows a first-order shift in oxidative cycling of Cr around 800 million years ago. Although we are working to refine the precise oxygen levels needed to induce oxidative Cr cycling in typical continental environments (see Project Report for Alternative Earth 1), these data indicate that prior to 800 million years ago, atmospheric pO2 may have been < 1% of the present atmospheric level (PAL), which is close to theoretical estimates for the minimum oxygen needs of the earliest metazoans (see Figure 1). If correct, this is strong evidence that environmental oxygen levels were low enough to have directly impacted the diversification of complex life, a premise that a number of researchers have challenged in the last few years. In fact, it is possible that oxygen levels were lower for periods of the Proterozoic than they were during transient oxygen highs in the Archean. If correct, this conclusion marks one of the most fundamental shifts in our view of Earth’s oxygenation in decades—and provides strong impetus to continue our reassessment of basic aspects of Earth’s oxygen cycle and its relationship with co-evolving life and atmospheric compositions.
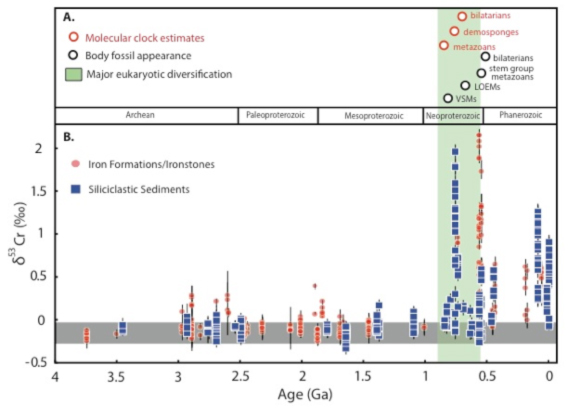
The chromium isotope proxy is the heart of our developing framework for very low atmospheric oxygen levels in the mid-Proterozoic; other independent proxies are indicating low oxygen in the shallow ocean and deep ocean as well (see Project Report for Alternative Earth 3). The importance of a full understanding of this pervasive oxygen stasis is difficult to overstate. The evolution of O2 levels in the mid-Proterozoic ocean-atmosphere system forms the backdrop for the initial emergence and subsequent evolutionary stasis of eukaryotic life—as recorded in the paleontological and organic geochemical records. Furthermore, the Cr data provide the possibility of a remarkably long period of Earth’s history during which many of the links among tectonics, climate, and life may have been short-circuited and/or amplified in unusual ways. For example, very low pO2 during mid-Proterozoic time likely limited O2 to “oxygen oases” within the Proterozoic ocean, and preliminary results suggest these oases—dynamic environments on biologically relevant timescales—were probably inhospitable for the origin of animal life despite recent suggestions for their low initial oxygen needs. If we are correct, this is a major conclusion that may explain the absence of animals during the mid-Proterozoic and the generally staid progress among all complex life.
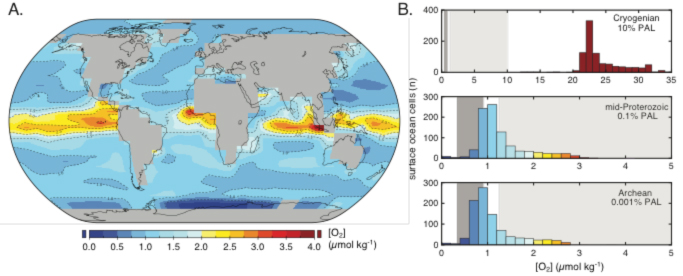
Inhospitable is a relative term, of course. No matter how stifling the mid-Proterozoic environment may have been compared to the modern Earth, it was still unequivocally inhabited. If an exo-world were similarly inhabited, we would like to know how to detect it. So, did that Earthly inhabitation produce a clear signal? Our modeling of biosignature gases during this time interval is revealing critical implications. In particular, recent isotopic constraints on evolving atmospheric pO2 and oceanic SO42- suggest the possibility that a range of conventional biosignature gases (O2, O3, CH4, N2O) would have been difficult to detect spectroscopically for much of mid-Proterozoic time, despite the great abundance of at least prokaryotic life in the ocean at that time (see Figure 2). These efforts highlight the importance of long-term chemical disequilibrium between the surface ocean and the atmosphere. This disequilibrium allows for the recycling of biosignature gases within the marine environment without detectable expression in the atmosphere.
In short, a critical implication of our mid-Proterozoic atmospheric modeling effort is that extended intervals of Earth history might have appeared sterile through the filter of sea-air exchange-assuming searches that rely on the current suite of proposed biosignature gases for remote life detection and currently available and planned telescope technologies. Our ongoing work seeks to identify novel biosignatures that are less likely to be muted by exchange with a liquid ocean, with the aspiration that these results will inform the design of future telescopes intended for exoplanetary life detection.
Interdisplinary Collaboration
We think of Earth’s atmosphere as the remotely detectable sum product of all the controlling factors on and within Earth that at any given time combine to sustain life. Those evolving compositions also capture evidence of great challenges to life—as during global-scale glaciations, major impact events, or severe nutrient limitation—and of life-sustaining tectonic processes. The primary objectives of our four Alternative Earths projects define a template for a broadly relevant and novel view of evolving redox and life on Earth, with the net expectation of resolving emergent atmospheric compositions that would bear the collective signature of the coupled tectonic and biogeochemical processes that produced them. To this end, our research strategy is carried out within four working groups, each broadly integrated in the range of expertise represented—with each informing, and being informed by, the others’ efforts.
Quite simply, we have been aiming since the inception of our proposal for an unusually, if not uniquely, unified effort that would not be possible outside the team structure of the NAI. Indeed, the whole already seems much greater than the sum of the parts. Our work on biotic versus abiotic cycling of N2O illustrates our team’s reliance on interdisciplinarity. In the lab, we are constraining the kinetics of N2O production in reducing abiotic systems, and then we leverage these experimental data in the context of atmospheric modeling—bringing together expertise at GT, UCR, and NASA’s Virtual Planetary Laboratory. The results of this work are already yielding important constraints on evolving climate system stability and the emergence and maintenance of remotely detectable atmospheric biosignatures during early periods of Earth’s history (see Project Report for Alternative Earth 2).
Our ongoing work on zinc (Zn) isotopes is another example of a truly interdisciplinary project that is only possible within a large collaborative network like our Alternative Earths Working Groups. Specifically, we are using Zn isotopes to track the rise of eukaryotes to ecological dominance and, at the same time, bridging those data with novel proteomics and quantitative modeling. In doing so, we have generated a new Zn isotope record that demonstrates that eukaryotes—although evolving before 1700 million years ago—only become important primary producers around 800 million years ago. These data provide a new view of cause-and-effect relationship between the diversification of complex life and evolution of Earth’s atmospheric composition (see Project Report for Alternative Earth 3).
Our Alternative Earths Team aims for a unified effort that would not be possible outside the structure of the NAI. Through collaboration among team members at GT, OHSU, Yale, and UCR, for example, we are studying the biogeochemistry of enzymatic manganese oxide formation and fractionation of stable isotopes under varying environmental conditions. Indeed, we brought together Yuanzhi Tang (GT) and Brad Tebo (OHSU) for this specific purpose. They had never met or collaborated before our Alternative Earths Team Kick-Off Meeting in April 2015 but have hit the ground sprinting since, including student exchanges and pursuit of independent funding for related projects (see Project Report for Alternative Earth 3).
Our team is also capitalizing on obvious synergism with the Origins of Complexity Team and the Virtual Planetary Laboratory (VPL). We have forged direct funded links to Origins of Complexity team member Greg Fournier (MIT) and to VPL team member Eddie Schwieterman through their 2015 NAI Director’s Discretionary Funding projects. Schwieterman’s plan is to couple our growing, comprehensive library of proxy-constrained estimates of ocean-atmosphere chemistry from the Proterozoic with the photochemistry/climate and radiative transfer models of the VPL. The ultimate goals are the integrated tools necessary to extend these diverse biogeochemical states toward a more complete understanding of a universe of possible exobiospheres. Fournier will study the rates and patterns of the acquisition and loss of genes coding for oxygen-associated enzymes in diverse microbial groups against a backdrop of changing biospheric oxygen as constrained by our team. The resulting insights into oxygenation dynamics during the Proterozoic should feed directly into our atmospheric modeling.
Collaboration between the Alternative Earths and Origins of Complexity NAI teams is bolstered by having members of both groups in the same department (Geology and Geophysics) at Yale. We have taken advantage of this opportunity by initiating several research efforts focused mainly on determining complex life’s role in shaping biogeochemical cycles and atmospheric evolution. (Indeed, we added a fourth Alternative Earth to our research repertoire based on our recent efforts and related ties to the Origins of Complexity (MIT) Team; our original proposal included only three Alternative Earths.) The productivity of this inter-team collaboration is already clear. Featuring authors from both teams, we recently published a paper in Nature Geoscience addressing how animals shaped the global sulfur and oxygen cycles—as well as an extensive review paper center on the interval leading up to the Snowball Earth glacial events. Furthermore, representatives from each team co-authored an article strengthening the case for early animal (sponge) biomarkers (see Project Report for Alternative Earth 4). In sum, we fully embrace the idea of inter-NAI team collaboration. We look forward to strengthening these ties as we move into Year 2 and beyond.
Publications
- Late Proterozoic transitions in climate, oxygen, and tectonics, and the rise of complex life. In: Earth-Life Transitions: Paleobiology in the Context of Earth System Evolution. N.J. Planavsky, L.G. Tarhan, E. Bellefroid, C.T. Reinhard, G. Love, T.W. Lyons, In P.D. Polly, J.J. Head, and D.L. Fox (eds.), Earth-Life Transitions: Paleobiology in the Context of Earth System Evolution. The Paleontological Society Papers 21. Yale Press, New Haven, CT.
-
Algeo, T. J., Luo, G. M., Song, H. Y., Lyons, T. W., & Canfield, D. E. (2015). Reconstruction of secular variation in seawater sulfate concentrations. Biogeosciences, 12(7), 2131–2151. doi:10.5194/bg-12-2131-2015
-
Glass, J. B. (2015). Microbes that Meddle with Metals. Microbe Magazine, 10(5), 197–202. doi:10.1128/microbe.10.197.1
-
Glass, J. B., Kretz, C. B., Ganesh, S., Ranjan, P., Seston, S. L., Buck, K. N., … Stewart, F. J. (2015). Meta-omic signatures of microbial metal and nitrogen cycling in marine oxygen minimum zones. Frontiers in Microbiology, 6. doi:10.3389/fmicb.2015.00998
-
Hansel, C. M., Ferdelman, T. G., & Tebo, B. M. (2015). Cryptic Cross-Linkages Among Biogeochemical Cycles: Novel Insights from Reactive Intermediates. ELEMENTS, 11(6), 409–414. doi:10.2113/gselements.11.6.409
-
Kendall, B., Komiya, T., Lyons, T. W., Bates, S. M., Gordon, G. W., Romaniello, S. J., … Anbar, A. D. (2015). Uranium and molybdenum isotope evidence for an episode of widespread ocean oxygenation during the late Ediacaran Period. Geochimica et Cosmochimica Acta, 156, 173–193. doi:10.1016/j.gca.2015.02.025
-
Lee, C., Love, G. D., Fischer, W. W., Grotzinger, J. P., & Halverson, G. P. (2015). Marine organic matter cycling during the Ediacaran Shuram excursion. Geology, None, G37236.1. doi:10.1130/g37236.1
-
Li, C., Planavsky, N. J., Love, G. D., Reinhard, C. T., Hardisty, D., Feng, L., … Lyons, T. W. (2015). Marine redox conditions in the middle Proterozoic ocean and isotopic constraints on authigenic carbonate formation: Insights from the Chuanlinggou Formation, Yanshan Basin, North China. Geochimica et Cosmochimica Acta, 150, 90–105. doi:10.1016/j.gca.2014.12.005
-
Li, C., Planavsky, N. J., Shi, W., Zhang, Z., Zhou, C., Cheng, M., … Xie, S. (2015). Ediacaran Marine Redox Heterogeneity and Early Animal Ecosystems. Scientific Reports, 5, 17097. doi:10.1038/srep17097
-
Love, G. D., & Summons, R. E. (2015). The molecular record of Cryogenian sponges – a response to Antcliffe (2013). Palaeontology, 58(6), 1131–1136. doi:10.1111/pala.12196
-
Luo, G., Ono, S., Huang, J., Algeo, T. J., Li, C., Zhou, L., … Xie, S. (2015). Decline in oceanic sulfate levels during the early Mesoproterozoic. Precambrian Research, 258, 36–47. doi:10.1016/j.precamres.2014.12.014
-
Lyons, T. W., Fike, D. A., & Zerkle, A. (2015). Emerging Biogeochemical Views of Earth’s Ancient Microbial Worlds. ELEMENTS, 11(6), 415–421. doi:10.2113/gselements.11.6.415
-
Osburn, M. R., Owens, J., Bergmann, K. D., Lyons, T. W., & Grotzinger, J. P. (2015). Dynamic changes in sulfate sulfur isotopes preceding the Ediacaran Shuram Excursion. Geochimica et Cosmochimica Acta, 170, 204–224. doi:10.1016/j.gca.2015.07.039
-
Partin, C. A., Bekker, A., Planavsky, N. J., & Lyons, T. W. (2015). Euxinic conditions recorded in the ca. 1.93Ga Bravo Lake Formation, Nunavut (Canada): Implications for oceanic redox evolution. Chemical Geology, 417, 148–162. doi:10.1016/j.chemgeo.2015.09.004
-
Stüeken, E. E., Buick, R., Bekker, A., Catling, D., Foriel, J., Guy, B. M., … Poulton, S. W. (2015). The evolution of the global selenium cycle: Secular trends in Se isotopes and abundances. Geochimica et Cosmochimica Acta, 162, 109–125. doi:10.1016/j.gca.2015.04.033
-
Thomson, D., Rainbird, R. H., Planavsky, N., Lyons, T. W., & Bekker, A. (2015). Chemostratigraphy of the Shaler Supergroup, Victoria Island, NW Canada: A record of ocean composition prior to the Cryogenian glaciations. Precambrian Research, 263, 232–245. doi:10.1016/j.precamres.2015.02.007
-
Glass, J. B., Kretz, C. B., Warren, M. J., & Ting, C. S. (n.d.). 7 Current perspectives on microbial strategies for survival under extreme nutrient starvation: evolution and ecophysiology. Microbial Evolution under Extreme Conditions. doi:10.1515/9783110340716.127