2015 Annual Science Report
 Massachusetts Institute of Technology
Reporting | JAN 2015 – DEC 2015
Massachusetts Institute of Technology
Reporting | JAN 2015 – DEC 2015
Early Animals: Modeling the Biotic-Abiotic Interface in the Early Evolution of Multicellular Form
Project Summary
Multicellular organisms in the sea modify their local hydraulic environment. Modeling of the earliest-known multicellular communities of frond-like forms demonstrated that they were large enough and closely spaced enough to generate a distinctive canopy flow-regime. In this context diffusion at the surface of organism was limiting and height and attendant velocity exposure permitted escape from these limits (Ghisalberti et al. 2014). Building on these results, we are developing models of abiotic/biotic interactions at organismal surfaces, relevant to the morphology, development and orientation of other Neoproterozoic fossils. A subset of these are flat-lying forms such as Dickinsonia. These may interact with the sediment modifying redox gradients. Ultimately, this work will help illuminate how forms initially dependent on passive diffusion became more trophically, morphologically and behaviorally complex, during the diversification of animals.
Project Progress
Our initial work on Precambrian communities of frond-like forms (Ghisalberti et al. 2014) established that given an assumption of uptake of reactants from the water as a mode of life (Osmotrophy) the hydraulic “canopy” developed by the community was critical to an advantage of vertical growth in these low-flow environments. We have now turned our attention to developing models of the hydraulic implications of flat-lying Ediacaran forms such as Dickinsonia.
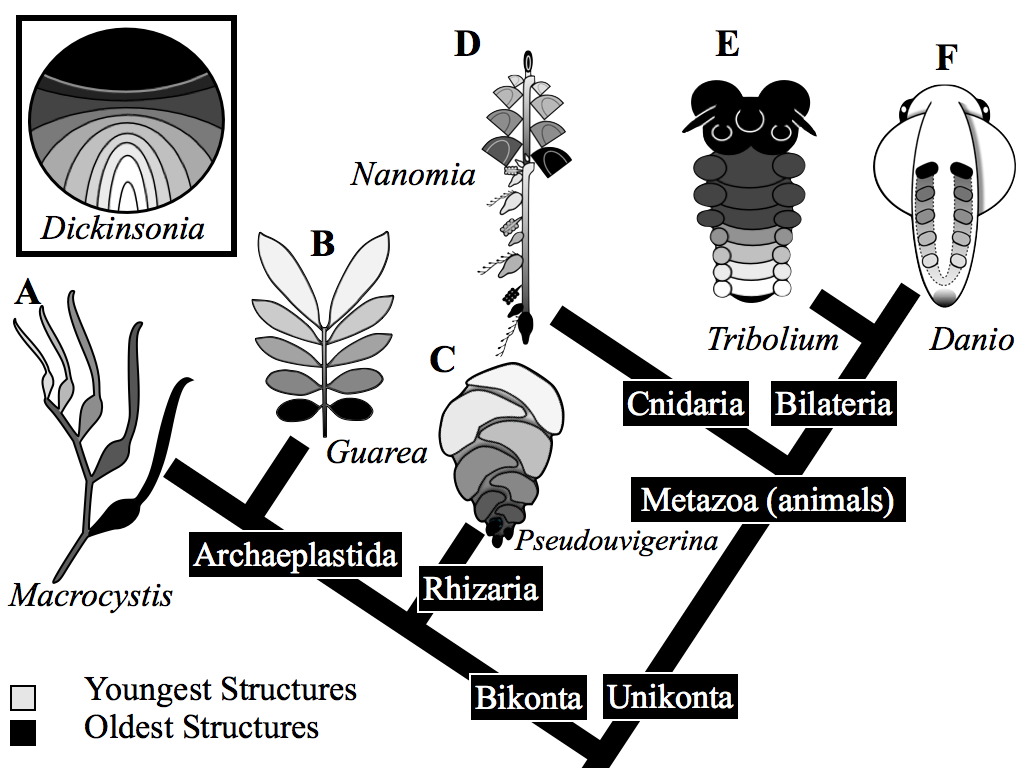
To provide an overall context for the work we have published an ancestral state reconstruction model linking the development process observed in Dickinsonia with the terminal addition process thought to be ancestral in Bilateria (see Fig. 1). This ancestral state reconstruction, based on development, provides an objective way to treat an otherwise enigmatic relationship of this flat-lying form to animal groups. The best supported inference, in this context, is that Dickinsonia shares a mode of development with Bilaterians.
Returning to hydraulic considerations, we anticipate that thin “epithelial” forms such as Dickinsonia, when placed in contact with the sediment surface, will modify redox gradients due to differences in permeability, and transport properties of biotic epithelial surface compared to the underlying sediment. This will change the sediment water column interaction and alter the redox profile, potentially to the benefit of the organism. We are currently developing simple one dimensional and two dimensional models that subsume attributes of diffusion and advection with and without an epithelial limit. We anticipate that this combined approach—where sedimentary environment, water movement and epithelia are modeled in concert—will illuminate aspects of the early evolution of multicellular form. Ultimately this modelling should provide an avenue to better model benthic-surface processes including those associated with mat settings in the deep-sea and those preserved in the rock record of early Earth.
This flow-modeling of the Ediacaran fauna with our colleague Marco Ghisalberti in Perth, is based on collaborative interactions established through the NASA astrobiology program. These were initiated following a NASA astrobiology field trip to Newfoundland and would not have developed without this NAI stimulus.
Additional work in the lab of potential interest to NASA includes study of the far-field effects of glaciation on biological evolution on coasts far from glacial ice. Other work in the lab relates climate, hydrology and estuarine and lagoonal biodiversity, and should allow us to predict climate vulnerabilities of species that inhabit these settings.
Publications
-
Gold, D. A., Runnegar, B., Gehling, J. G., & Jacobs, D. K. (2015). Ancestral state reconstruction of ontogeny supports a bilaterian affinity forDickinsonia. Evolution & Development, 17(6), 315–324. doi:10.1111/ede.12168
-
PROJECT INVESTIGATORS:
-
PROJECT MEMBERS:
Marco Ghisalberti
Co-Investigator
David Gold
Co-Investigator
David Johnston
Co-Investigator
Marc LaFlamme
Co-Investigator
Roger Summons
Co-Investigator
Tina Treude
Collaborator
-
RELATED OBJECTIVES:
Objective 4.1
Earth's early biosphere.
Objective 4.2
Production of complex life.
Objective 5.2
Co-evolution of microbial communities
Objective 6.1
Effects of environmental changes on microbial ecosystems
