2013 Annual Science Report
 University of Hawaii, Manoa
Reporting | SEP 2012 – AUG 2013
University of Hawaii, Manoa
Reporting | SEP 2012 – AUG 2013
Habitability, Biosignatures, and Intelligence
Project Summary
Understanding the nature and distribution of habitable environments in the Universe is one of the primary goals of astrobiology. Based on the only example of life we know, we have devel-oped various concepts to predict, detect, and investigate habitability, biosignatures and intelli-gence occurrence in the near-solar environment. In particular, we are searching for water vapor in atmospheres of extrasolar planets and protoplanets, developing techniques for remote detec-tion of photosynthetic organisms on other planets, have detected a possible bio-chemistry sig-nature in Martian clays contemporary with early life on Earth, developed a comprehensive methodology and an interactive website for calculating habitable zones in binary stellar systems, expanded on definitions of habitable zones in the Milky way Galaxy, and proposed a novel ap-proach for searching extraterrestrial intelligence.
Project Progress
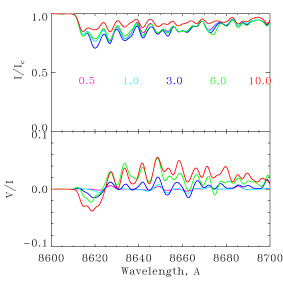
A search for water and other molecules in exoplanetary atmospheres [Berdyugina]
To clarify how planets acquire liquid water and how planet formation influences their environments and habitability, we investigate hot water (T>300K) and other molecules in protoplanets and ex-oplanets irradiated by the central star. Our goals are to determine what is the amount of water in planetesimals in protoplanetary disks and in the asteroid belt of the Solar system, what is the range of atmospheric conditions on planets with water, and how magnetic fields evolve in planets and shield planetary atmospheres from erosion by the stellar wind. To achieve these goals, we have further advanced the theory of scattering molecular polarization in thin atmospheres. This theory is used to synthesize spectra from upper planetary atmospheres (above a cloud deck), cometary comae, and exospheres of ice-rich asteroids which are remnants of planet formation in the Solar system. We have obtained time series of broad-band polarmetric measurements of hot Jupiters to determine their geometrical albedo and colors, first spectropolarimetric observations of exoplanets orbiting nearby stars, the dwarf planet Ceres, which is the closest to Earth protoplanet, and the Saturn moon Enceladus, which has now an active geiser pooring water from unbeneath its ice crust into the space. We are analysing these data to detect scattering polarization in water bands and determine limits on its abundance in these objects. We have also developed a novel technique to detect magnetic fields on brown dwarfs and hot Jupiters using CrH molecule transitions in the near infrared (see Figure 1). We have obtained spectropolarimetric measurements of brown dwarfs and investigate their magnetism. This opens the door to potential direct measurements of magnetic fieldsnd on hot Jupiters and investigate their interaction with stellar wind and acticity.
Habitable zones in planet-hosting binary star systems [Haghighipour]
We have developed a comprehensive methodology for calculating the boundaries of the habitable zone (HZ) of planet-hosting binary star systems. Our approach is general and takes into account the contribution of each star of the binary to the total flux received at the top of the atmosphere of an Earth-like planet, based on the star’s spectral energy distribution. We have studied how the binary eccentricity and stellar energy distribution affect the extent of the habitable zone. We have applied our calculations to binaries consisting of solar-type and red dwarf stars and inter-binary planets, and also to two known such planet-hosting binaries alpha Cen AB and HD 196886. We have also developed similar methodology for binary systems with circumbinary planets, and applied our calculations to known Kepler’s planetary systems (see Figure 2). We show that the HZ in binary systems is dynamic and depending on the luminosity of the binary stars, their spectral types, and the binary eccentricity, its boundaries vary as the stars of the binary undergo their orbital motion. We have developed a website where we show the results of the calculations of the HZ, along with movies of their dynamical evolution. The website can be found at http://astro.twam.info/hzptype.
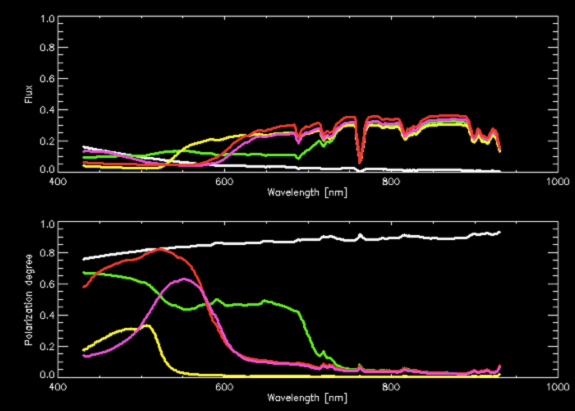
Detecting biosignatures of Earth-like planets in habitable zones with polarimetry [Berdyugina]
Life on Earth is aware of light polarization and makes good use of it for its survival and growth. For example, scattering polarization of the sky provides reliable directions, while specular reflection from water and plants helps life find healthy environments. Plants were found to grow faster if illuminated by left-handed circularly polarized light, and insects use polarized light to recognize their mates. In all cases, it is the solar light that is reflected, processed and analyzed by life forms, and the same circumstances are expected to exist on all habitable planets. Photosynthesis, in particular, is very likely to arise on another planet and can produce conspicuous biosignatures. We have pro-posed a novel remote-sensing method for detecting and identifying life forms on distant worlds and distinguishing them from non-biological processes. To achieve this we have designed and built a bio-polarimetric laboratory experiment for measuring optical polarized spectra of various biological and non-biological samples. We have focused on photosynthetic life molecules – pigments, which are common in plants, algae and bacteria. Photosynthesis is very likely to emerge early and last long on other planets with life. Moreover, we expect that the basic structure of photosynthetic molecules will be similar to that of terrestrial samples, as their main function is to capture photons and store their energy in chemical bonds. We have carried out laboratory spectropolarimetric measurements of a representative sample of plants containing various amounts of pigments such as chlorophyll, carotenoids, and anthocyanins. We have also measured a variety of non-biological samples (sands, rocks). Using our lab measurements, we have modeled intensity and polarized spectra of Earth-like planets having different surface coverage by photosynthetic organisms, deserted land, and ocean, as well as clouds (Figure 3). Our results demonstrate that polarized spectra provide very sensitive and unambiguous detection of photosynthetic organisms of various kinds. Our work paves the path towards analogous measurements of microorganisms and remote sensing of microbial ecology on the Earth and of extraterrestrial life on other planets and moons.
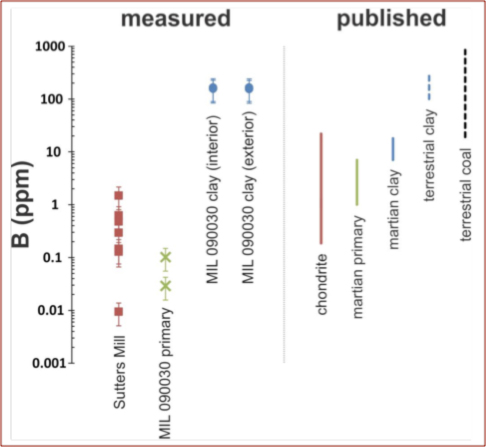
Boron enrichment in Martian clay [Hallis & Stephenson]
We have determined the abundance of boron in the alteration veins of Martian meteorite MIL 090030, via in-situ Cameca ims 1280 ionmicroprobe analyses. For comparison we also analyzed the primary igneous minerals of MIL 090030, as well as various primary and aqueous alteration phases in Sutters Mill – a carbonaceous chondrite meteorite observed to fall in 2012. We found that boron is concentrated in martian alteration veins (~160 ppm), to levels rivaling those found in terrestrial clays and marine sediments (see Figure 4). The level of boron concentration in the alteration veins of MIL 090030 cannot be explained by terrestrial contamination, as no atmospheric reservoir on Earth approaches the abundance of boron measured in these alteration veins. Also, both an exterior and interior sample from the MIL 090030 meteorite stone were measured. We found no measureable difference between boron abundances in the alteration veins of the internal and external areas, implying terrestrial boron is negligible and that the boron concentration in MIL 090030 alteration veins must be the result of secondary alteration processes on Mars.
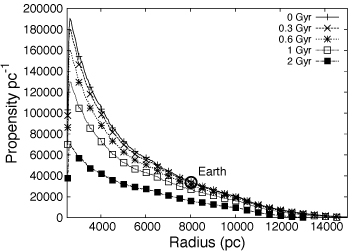
Galactic habitable zone and the propensity for the emergence of intelligent life [Gowanlock, Gazan]
The habitability of the Milky Way galaxy has been a subject of a recent study as a combined result of exoplanet detections and numerous conditions that life is found to thrive on the Earth. In our previous work of Gowanlock et al. (2011), we studied the habitability of the Milky Way by considering some of the prerequisites for complex life, such as having enough metallicity to form habitable planets, enough time for biological evolution and considering the negative effects of supernovae sterilizations. In our previous work, the timescales required for complex life to develop assumes the timescales for the development of land-based metazoa on Earth. Based on timescales taken from the origin and evolution of life on Earth, the model suggests large numbers of potentially habitable planets may exist in our Galaxy, with the greatest concentration being located towards the inner galaxy at R>2.5 kpc. Moving forward with this research, we are working to address the propensity for the emergence of intelligent life in the Milky Way. We utilize the planets and associated sterilization histories modelled in Gowanlock et al. (2011) to estimate the propensity for the emergence of intelligent life. The planets in the model form at a time consistent with an inside-out formation history of the Milky Way. We assume that the rise of intelligent life is influenced by the amount of time available for the evolutionary process to proceed without resetting (sterilization) events. We analyze the timing of supernovae events and the time between these events that could provide an opportunity for the development of intelligent life. Across propensity metrics, we find that even if we require long time durations between sterilizations for the development of intelligent life, the inner Galaxy provides the greatest number of opportunities at R>2.5 kpc despite the high supernovae rate in the region (see preliminary result in Figure 5). The results suggest that the inner Galaxy is a good location for SETI searches.
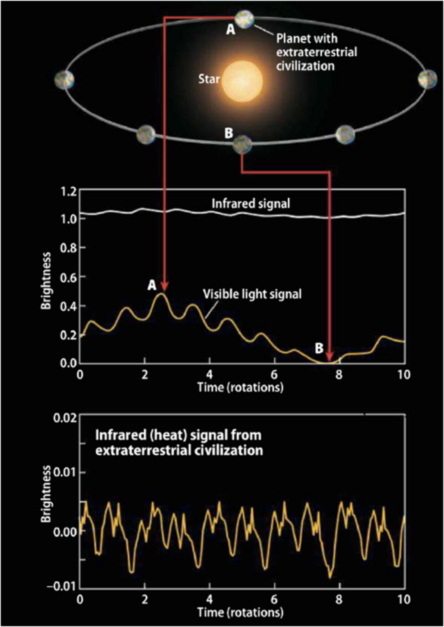
A search for extraterrestrial intelligent life with infrared light [Berdyugina]
For a half-century searches have been conducted at radio and optical frequencies for evidence of extraterrestrial civilizations (ETCs). These approaches depend on finding alien transmissions, beam “leakage”, or what could be called intentional electromagnetic signals from ETCs that are operating cosmic beacons. In this project, we have developed a strategy for detecting an unintentional signal caused by alien planetary warming. We have proposed a dual wavelength, visible and infrared, search (see Figure 6) that would be largely independent of alien communication modes and would have quantifiable statistical completeness. We have shown that it is possible to detect an unintentional thermodynamic signal from Earth-like ETC’s within 60 light years from the Sun with current technology. Even a null result would help us understand the Fermi paradox, “why do we appear to be alone?” The detection of an alien heat signature, even without further communication, will be important – if only to demonstrate the possibility of civilization achieving a phase of sustainable global-scale power consumption.

Publications
-
Eggl, S., Haghighipour, N., & Pilat-Lohinger, E. (2013). DETECTABILITY OF EARTH-LIKE PLANETS IN CIRCUMSTELLAR HABITABLE ZONES OF BINARY STAR SYSTEMS WITH SUN-LIKE COMPONENTS. The Astrophysical Journal, 764(2), 130. doi:10.1088/0004-637x/764/2/130
-
Eggl, S., Pilat-Lohinger, E., Funk, B., Georgakarakos, N., & Haghighipour, N. (2012). Circumstellar habitable zones of binary-star systems in the solar neighbourhood. Monthly Notices of the Royal Astronomical Society, 428(4), 3104–3113. doi:10.1093/mnras/sts257
-
Gowanlock, M. G., Patton, D. R., & McConnell, S. M. (2011). A Model of Habitability Within the Milky Way Galaxy. Astrobiology, 11(9), 855–873. doi:10.1089/ast.2010.0555
-
Haghighipour, N. (2013). The Formation and Dynamics of Super-Earth Planets. Annual Review of Earth and Planetary Sciences, 41(1), 469–495. doi:10.1146/annurev-earth-042711-105340
-
Haghighipour, N., & Kaltenegger, L. (2013). CALCULATING THE HABITABLE ZONE OF BINARY STAR SYSTEMS. II. P-TYPE BINARIES. The Astrophysical Journal, 777(2), 166. doi:10.1088/0004-637x/777/2/166
-
Kaltenegger, L., & Haghighipour, N. (2013). CALCULATING THE HABITABLE ZONE OF BINARY STAR SYSTEMS. I. S-TYPE BINARIES. The Astrophysical Journal, 777(2), 165. doi:10.1088/0004-637x/777/2/165
-
Kuzmychov, O., & Berdyugina, S. V. (2013). Paschen-Back effect in the CrH molecule and its application for magnetic field measurements on stars, brown dwarfs, and hot exoplanets. A&A, 558, A120. doi:10.1051/0004-6361/201220041
-
Melchiorre, E. B., Huss, G. R., & Lopez, A. (2014). Carbon and hydrogen stable isotope microanalysis and data correction for rare carbonate minerals: Case studies for stichtite (Mg6Cr2[(OH)16|CO3]∙H2O) and malachite (Cu2CO3(OH)2). Chemical Geology, 367, 63–69. doi:10.1016/j.chemgeo.2013.11.010
-
Müller, T. W. A., & Haghighipour, N. (2014). CALCULATING THE HABITABLE ZONES OF MULTIPLE STAR SYSTEMS WITH A NEW INTERACTIVE WEB SITE. The Astrophysical Journal, 782(1), 26. doi:10.1088/0004-637x/782/1/26
-
Stephenson, J. D., Hallis, L. J., Nagashima, K., & Freeland, S. J. (2013). Boron Enrichment in Martian Clay. PLoS ONE, 8(6), e64624. doi:10.1371/journal.pone.0064624
- Berdyugina, S.V., Berdyugin, A.V. & Piirola, V. (2012). Albedo of exoplanets constrained by polarimetry. In: Hoffman, J. (Eds.). Polarimetry: From Birth to Death. Vol. 1429.
- Berdyugina, S.V., Berdyugin, A.V. & Piirola, V. (2013). Upsilon Andromedae b in polarized light: New constraints on planet size, density, and albedo. Astronomy & Astrophysics (A&A).
- Kuhn, J.R., Berdyugina, S.V., Halliday, D. & Harlingten, C. (2013). How to find ET with Infrared Light, Astronomy. Astronomy.
- Kuzmychov, O. & Berdyugina, S.V. (2012). CrH molecule: A new diagnostic tool for measuring magnetic fields of ultra-cool dwarfs. In: Hoffman, J. (Eds.). Stellar Polarimetry: From Birth to Death. Vol. 1429.
- Kuzmychov, O. & Berdyugina, S.V. (2013). CrH molecule: New diagnostic tool for measuring magnetic fields of cool dwarfs [Book Chapter]. Solar and Astrophysical Dynamos and Magnetic Activity. Vol. 294.
- Kuzmychov, O., Berdyugina, S., Harrington, D. & Kuhn, J. (2013). Investigating the magnetism of brown dwarfs, in Brown Dwarfs. Memorie della Societa Astronomica Italiana, 84(1127).
-
PROJECT INVESTIGATORS:
-
PROJECT MEMBERS:
Svetlana Berdyugina
Project Investigator
Stephen Freeland
Co-Investigator
Nader Haghighipour
Co-Investigator
Lydia Hallis
Co-Investigator
David Harrington
Co-Investigator
Lisa Kaltenegger
Co-Investigator
Jeffrey Kuhn
Co-Investigator
James Stephenson
Co-Investigator
Mike Gowanlock
Collaborator
Kazuhide Nagashima
Collaborator
-
RELATED OBJECTIVES:
Objective 1.1
Formation and evolution of habitable planets.
Objective 1.2
Indirect and direct astronomical observations of extrasolar habitable planets.
Objective 2.2
Outer Solar System exploration
Objective 3.1
Sources of prebiotic materials and catalysts
Objective 3.2
Origins and evolution of functional biomolecules
Objective 4.1
Earth's early biosphere.
Objective 4.2
Production of complex life.
Objective 6.2
Adaptation and evolution of life beyond Earth
Objective 7.1
Biosignatures to be sought in Solar System materials
Objective 7.2
Biosignatures to be sought in nearby planetary systems