2012 Annual Science Report
 University of Wisconsin
Reporting | SEP 2011 – AUG 2012
University of Wisconsin
Reporting | SEP 2011 – AUG 2012
Project 1A: Interaction Between Lipid Membranes and Mineral Surfaces
Project Summary
The goal of our work was to determine whether the surface chemistry or physical properties of mineral surfaces in contact with model “protocells” may have controlled the rupture/integrity of the protocell membranes. We hypothesized that mineral surface/lipid head group interface chemistry may have acted as a Darwinian selection force for the types of protocells which survived. We used a variety of experimental techniques including fluorimetric analysis of vesicle rupture in contact with mineral particles, bulk adsorption isotherms for lipids on oxide particles, Atomic Force Micropscopy and Neutron Reflectivity studies of lipid bilayer formation on planar oxide surfaces. The results could be explained by a modified version of the Deraguin-Landau-Verwey-Ovebeek (DLVO) theory for colloidal stability. Results indicated that lipid vesicles and bilayers are more stable at the positively-charged corundum surface in the low and mid-pH range than at the negatively-charged surface at high pH. Thus, protocells which came in contact with mineral surfaces having a positive surface charge would have maintained their integrity while those contacting negatively-charged surfaces would have been ruptured. Our results may suggest that mineral surface chemistry and lipid head group chemistry may have acted as a Darwinian “evolutionary stress” to select the most robust types of vesicles which “survived.”
Our project addresses NASA Astrobiology Institute’s (NAI) Roadmap goals of understanding the origins of cellularity and the evolution of mechanisms for survival at environmental limits, and NASA’s Strategic Goal of advancing scientific knowledge of the origin and evolution of the Earth’s biosphere and the potential for life elsewhere.
Project Progress
The goal of our work was to determine whether the surface chemistry or physical properties of mineral surfaces in contact with model “protocells” may have controlled the rupture/integrity of the protocell membranes. We hypothesized that mineral surface/lipid head group interface chemistry may have acted as a Darwinian selection force for the types of protocells which survived. We used phospholipid vesicles to represent prebiotic protocells although we recognize that phospholipids are biologically produced, and that simpler lipids would have existed pre-biotically. Model mineral quartz (α-SiO2), corundum (α-Al2O3) and rutile (α -TiO2) particles (~ 1 mm diameter particles) were chosen to represent a range of surface charges, dielectric constants, and mineral solvation properties, and because the surface chemistry of these oxides is well-established in the literature. Physical properties, such as relative surface curvature of the model vesicle versus mineral, was accounted for by conducting additional studies using planar oxide supports. We chose lipid head groups of various charges, namely, zwitterionic phosphatidylcholine (DPPC or DTPC), anionic dipalmitoylphosphatidylserine (DPPS), and cationic dipalmitoylethylphosphatidylcholine(DPEPC).
We used a variety of experimental techniques including fluorimetric analysis of vesicle rupture in contact with mineral particles, bulk adsorption isotherms for lipids on oxide particles, Atomic Force Micropscopy and Neutron Reflectivity studies of lipid bilayer formation on planar oxide surfaces. The results could be explained by a modified version of the Deraguin-Landau-Verwey-Ovebeek (DLVO) theory for colloidal stability.
Using fluorimetric experiments on fluorescent dye-loaded DPPC vesicles in the presence of mineral particles, we determined that the greatest extent of DPPC vesicles was induced by quartz surfaces, followed by corundum and the least rupture was obtained on rutile (Figure 1) (Oleson and Sahai, 2010; Oleson et al., 2010). Furthermore, vesicle rupture occurred on quartz regardless of thermal cycling across the lipid gel–liquid crystal phase transition temperature, but rupture was promoted by cycling on corundum and rutile. If we consider vesicle rupture as a model process for lysis of protocell membranes, our results suggest that quartz has a greater ability to induce membranolysis than the other two mineral surfaces.
After rupturing in contact with all three mineral surfaces, the lipid molecules self-assembled at the surfaces into bilayers similar to model membranes (Oleson and Sahai, 2010; Oleson et al., 2010; Xu et al., 2009; Stevens, M.S. Thesis). The affinity of bilayers at mineral surfaces represents the protocell stability or “survivability”. Adsorption isotherms of DTPC at pH 7.2 in 0.017 M NaCl solution (above DTPC transition temperature) showed only two lipid bilayers on quartz compared to three on corundum and rutile (Figure 2). These results, again, indicated a lower affinity of the PC bilayers on the quartz surface, which is negatively-charged and slightly less hydrophilic than the other two mineral surfaces. The changes in the number of bilayers obtained at various pHs, and at various ionic strengths set by NaCl, with or without Ca2+ (Figure 3) were explained by our modified Deraguin-Landau-Verwey-Ovebeek (DLVO) theory for colloidal stability (Figure 4). According to this theory, the interaction energy between the oxide surface and the lipid bilayers depends on electrostatic forces and van der Waals forces. If DPPC bilayers were assumed to bear a small negative potential, then van der Waals forces predominantly accounted for two DPPC bilayers, and adsorption beyond the second bilayer occurred at low ionic strength due to extension of the electric double-layer near the oxide surface. Inclusion of bilayer–bilayer interactions resulted in a prediction of infinite DPPC bilayer deposition irrespective of the oxide. This was not observed experimentally, perhaps, due to the systems reaching quasi steady-state conditions rather than equilibrium. In contrast, adsorption isotherms of anionic DPPS and cationic DPEPC lipids show that adsorption of highly charged bilayers was decreased or prevented altogether due to bilayer-oxide and/or bilayer–bilayer repulsion (Figure 3).
We confirmed the greater affinity of corundum for PC head-groups using TappingMode Atomic Force Microscopy (AFM)to examine bilayers of ditridecanoylphosphatidylcholine, (DTPC; Tm = 14 oC) formed on planer surfaces of quartz and corundum at room temperature (Xu et al., 2009) (Figure 5) (Xu et al., 2009; Stevens, 2007, M.S. Thesis). Plan views and height profiles indicate the formation of incomplete single bilayer patches on quartz compared to at least two bilayers on corundum. Thus, the trends in lipid adsorption in this experimental set-up using planar oxide surfaces remain the same as in the adsorption isotherms where oxide particles were used: The lipids have a greater affinity for the positively-charged corundum surface compared to the negatively-charged quartz surface. Other oxides such as amorphous silica glass, rutile and muscovite mica also exhibited consistent trends of increasing DTPC adsorption with increasing positive surface charge of the mineral.
Neutron reflectivity studies of (DPPC) bilayers were conducted for the first time for bilayers deposited from solution on the (110) face of sapphire (corundum, α-Al2O3), at National Institute of Science and Technology (NIST). Bilayer coverage on planar sapphire depended on pH, ionic strength and Ca2+ (Figure 6) (Oleson et al., 2012). Surface coverage was higher at low to mid pH and lower ionic strength where the surface has a larger positive surface charge and is lower at higher pH and higher ionic strength or in the presence of Ca2+. These trends were consistent with previous adsorption isotherm results using corundum particles in aqueous suspensions and AFM studies on planar corundum surfaces.
In summary, mineral surface potential and relative hydrophilicity, lipid bilayer surface potential, and solution composition and temperature cycling, among other factors, could have controlled the self-assembly or lysis of early proto-cell membranes composed of amphiphilic lipids or lipid precursors such as fatty acids and fatty acid esters. Changes in solution conditions such as pH or ionic strength due to wetting–drying cycles in a tidal or lacustrine environment or due to a chemical gradient in seawater, for e.g., with distance from a submarine hydrothermal vent, could lead to different numbers of bilayers associated with mineral surfaces. Our results suggest that the self-assembly and stability of early proto-cell lipid membranes would have been favored on surfaces of minerals with positively-charged and more hydrophilic nature, and which were more abundant than the model corundum used here, compared to minerals with negatively-charged and less hydrophilic surfaces such as quartz or minerals of similar surface chemistry.
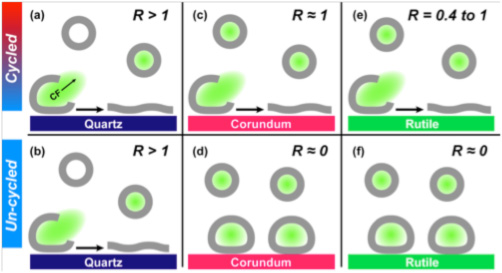
Effects of mineral surface chemistry and thermal cycling of lipid through the melting point on model protocell (DPPC vesicle) rupture or integrity. Schematic shows carboxyfluorescein (green)-loaded vesicles (grey outline) interacting with quartz (blue) and corundum (pink) surfaces which, respectively, have negative and positive surface charge.
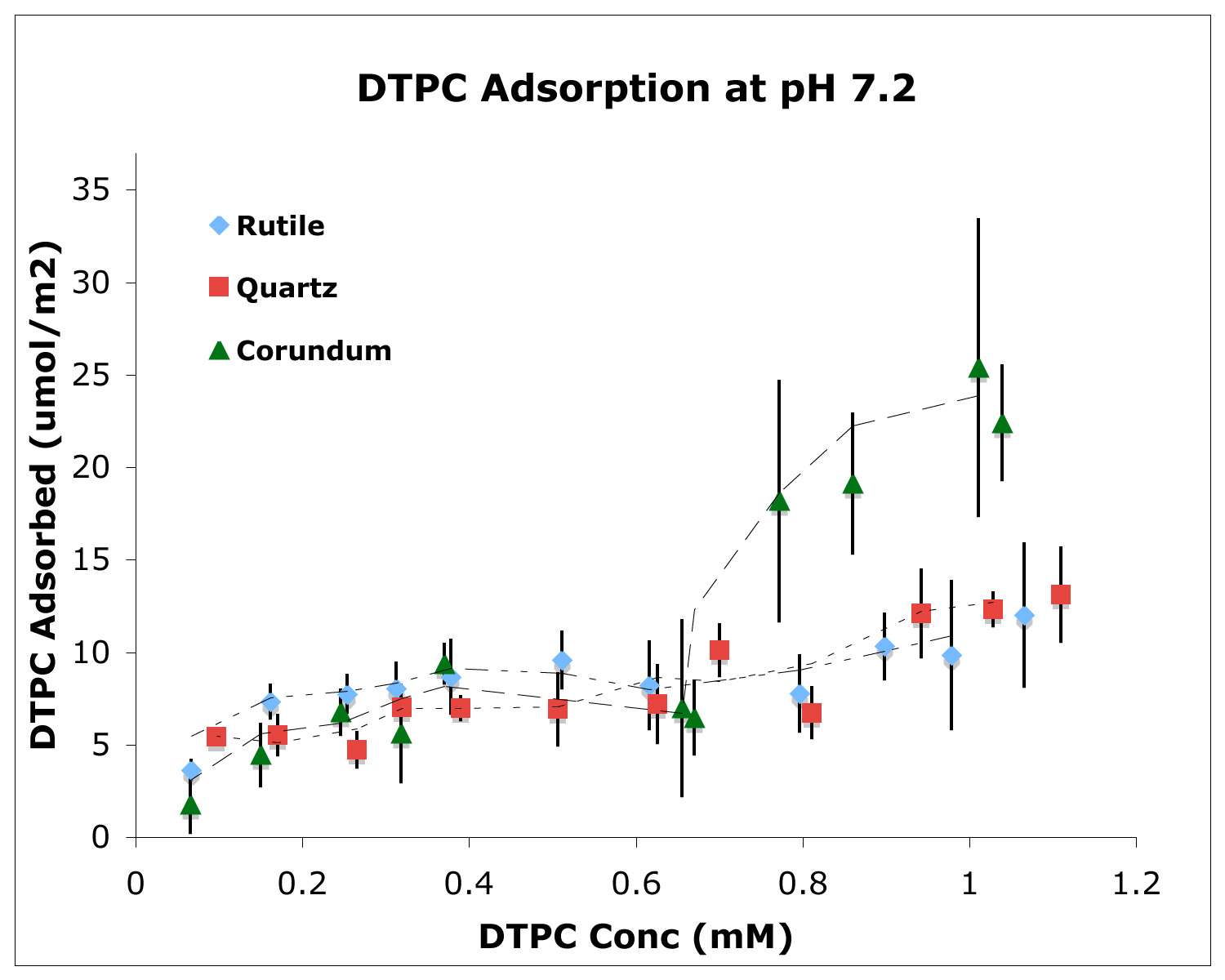
Adsorption isotherm of the phospholipid, DTPC, on micron-sized particulate quartz, rutile and corundum, which are chosen to represent different mineral surface charges. The isotherm plateaux for quartz and rutile correspond to two adsorbed lipid bilayers, whereas that for corundum represents three adsorbed bilayers.
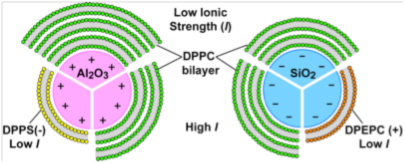
Schematic summarizing adsorption affinities of lipids having various surface charges (DPPC, DPPS, DPEPC) on mineral surfaces of different charges. The effects of ionic strength on lipid adsorption are also indicated.
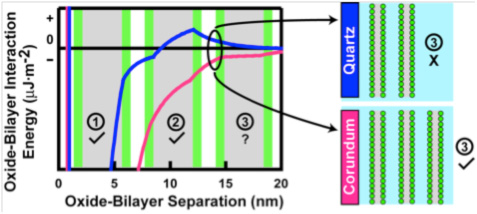
Modified DLVO theory predictions for adsorption of DPPC bilayers on quartz and corundum. Note that two bilayers are predicted for quartz and three for corundum, as obtained by the adsorption isotherms.
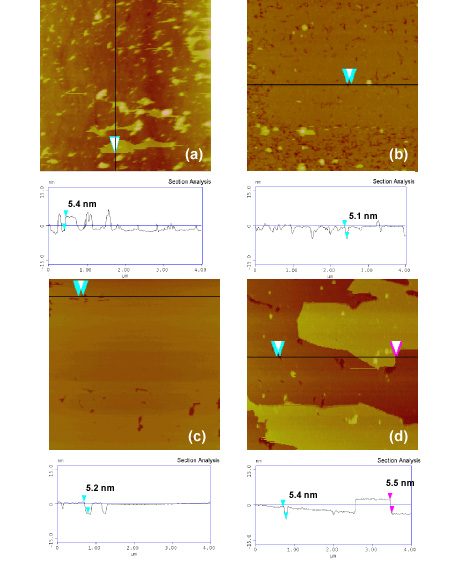
Atomic Force Microscopy (AFM) plan views and vertical cross-sections of lipid bilayers deposited on various planar oxide surfaces. Each bilayer is about 5-6 nm thick, and the images show a greater number of bilayers on corundum than on the other surfaces. The color scale represents vertical height above the surface, with lighter colors being higher and darker colors being lower.
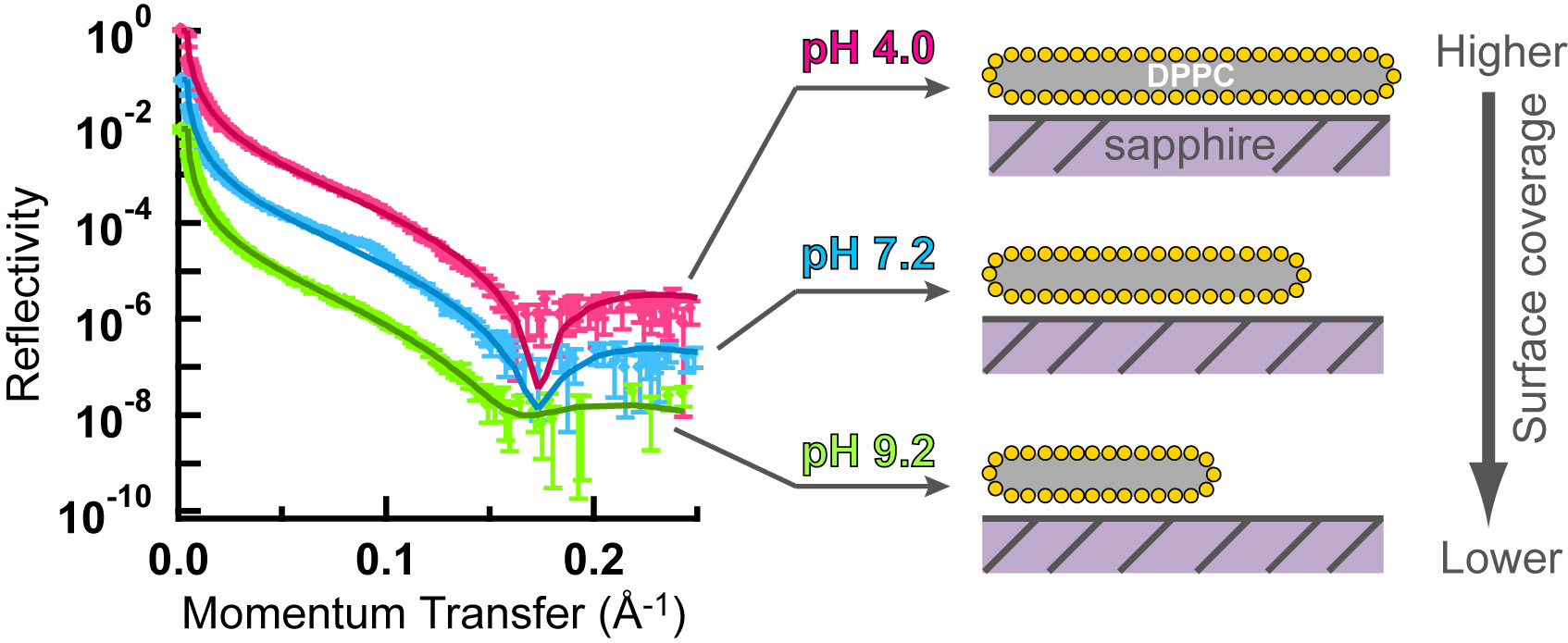
Schematic summarizing neutron reflectivity results for DTPC bilayer formation on planar corundum (sapphire) surface at various pHs. The corundum surface is positively-charged at low pHs and negatively-charged at high pHs. Results show greater lipid bilayer adsorption at the positively- charged surface than the negatively-charged surface, consistent with our results using various other techniques as shown above.
Publications
-
Oleson, T. A., & Sahai, N. (2010). Interaction energies between oxide surfaces and multiple phosphatidylcholine bilayers from extended-DLVO theory. Journal of Colloid and Interface Science, 352(2), 316–326. doi:10.1016/j.jcis.2010.08.056
-
Oleson, T. A., Sahai, N., & Pedersen, J. A. (2010). Electrostatic effects on deposition of multiple phospholipid bilayers at oxide surfaces. Journal of Colloid and Interface Science, 352(2), 327–336. doi:10.1016/j.jcis.2010.08.057
-
Oleson, T. A., Sahai, N., Wesolowski, D. J., Dura, J. A., Majkrzak, C. F., & Giuffre, A. J. (2012). Neutron reflectivity study of substrate surface chemistry effects on supported phospholipid bilayer formation on
-
Xu, J., Stevens, M. J., Oleson, T. A., Last, J. A., & Sahai, N. (2009). Role of Oxide Surface Chemistry and Phospholipid Phase on Adsorption and Self-Assembly: Isotherms and Atomic Force Microscopy. The Journal of Physical Chemistry C, 113(6), 2187–2196. doi:10.1021/jp807680d
- Oleson, T.A. & Sahai, N. (2008). Oxide-dependent adsorption and self-assembly of dipalymitoylphosphatidylcholine, a cell-membrane phospholipid: Bulk Adsorption Isotherms. Langmuir, 24: 4865-4873.
-
PROJECT INVESTIGATORS:
-
PROJECT MEMBERS:
Timothy Oleson
Graduate Student
Mark Stevens
Graduate Student
Jie Xu
Graduate Student
-
RELATED OBJECTIVES:
Objective 3.4
Origins of cellularity and protobiological systems
