2012 Annual Science Report
 Arizona State University
Reporting | SEP 2011 – AUG 2012
Arizona State University
Reporting | SEP 2011 – AUG 2012
Stoichiometry of Life, Task 3a: Ancient Records - Geologic
Project Summary
Fossil and chemical fingerprints of animal life first appear in the geologic record around 600 million years ago. The four billion years of Earth history before this milestone were marked by dramatic changes that we take for granted today but that set the stage for our existence. Among the key events recorded in very old rocks is the first rise of oxygen in the atmosphere and ocean about 2.5 billion years ago following two billion years of a virtually oxygen-free world. And this evolving chemical state was the backdrop against which photosynthesis first evolved; simple, single-celled organisms appeared and diversified; and the first eukaryotic life evolved as a forerunner to the complex animals that would follow one-to-two billion years later. Our work is exploring the evolving compositions of the early atmosphere and ocean and their cause-and-effect relationships with the evolution of life—spanning the middle 50% of Earth history from the first production of oxygen via photosynthesis to the first appearance of animals. Darwin would have been pleased to know that early rocks tell us a convincingly strong: long before the animals, the oceans were teeming with life and that this life set the stage, in so many ways, for the later evolution of animals. Our sophisticated geochemical tracers are changing our view of the early environmental conditions that facilitated, and just as often throttled, the rise of life and the ways life can passively and intentionally modify its own environment—not unlike the lessons we are learning about our relationship with the changing ocean, atmosphere, and climate today.
Project Progress
Building from our work over the past few years, we have concentrated our efforts over the last year on the evolving redox state of the atmosphere and ocean between roughly 3.0 and 0.6 billion years ago. This window spans for the earliest evidence for oxygenic photosynthesis to the first appearance of animals, and we have focused in particular on the mid-Proterozoic ‘boring billion’ noted for its persistence of oxygen-lean conditions in the deep ocean long after the first oxygenation of the atmosphere. Our work suggests that prevailing ocean redox during the mid-Proterozoic and coupled nutrient limitations may have throttled the proliferation of prokaryotic life, the early rise of eukaryotes, and ultimately the first appearance of animals-and, in the process, the burial of organic matter and the further rise of oxygen. We are most interested in the feedbacks that may have set the steady-state oxygen conditions at low, higher, and very high levels and when and why the biosphere stepped, not necessarily irreversible, between those levels.
To this end, the efforts of the past year have focused on:
(1) development of novel proxies (e.g., selenium and uranium; manuscript preparation and publication),
(2) calibration and testing of more established proxies (e.g., iron and molybdenum isotopes; manuscript publication),
(3) characterization of the redox state of the late Archean atmosphere and ocean and the transition across the Great Oxidation Event (manuscript preparation and publication),
(4) conditions in the late Neoproterozoic ocean and relationships to a rise in oxygen and the appearance of animals (final stage of data generation and interpretation, manuscript preparation and publication),
(5) Conditions in the mid-Proterozoic ocean (data interpretation, manuscript preparation and publication, extensive generation of new data). We have prioritized mid-Proterozoic rocks from N. Australia and N. China, including exhaustive generation of new organic and inorganic data (organic biomarker extraction/analysis, Fe speciation, trace metals, S isotope analysis, etc.) for the Roper Group in Australia. These data are speaking to the redox conditions in the deep ocean, nutrient availability, and, importantly, whether sulfidic conditions were widespread in the shallowest parts of the mid-Proterozoic ocean. Our organic data, when viewed in light of complementary inorganic and sedimentological results, suggest that shallow euxinia was rare and should not factor prominently in models for primary production for that ocean (via anoxygenic photosynthesis by S oxidizing bacteria). All our data are leading toward a unified view of the Precambrian ocean, including the likely persistence of anoxic, iron-rich deep waters for most of the first 90% of Earth history.
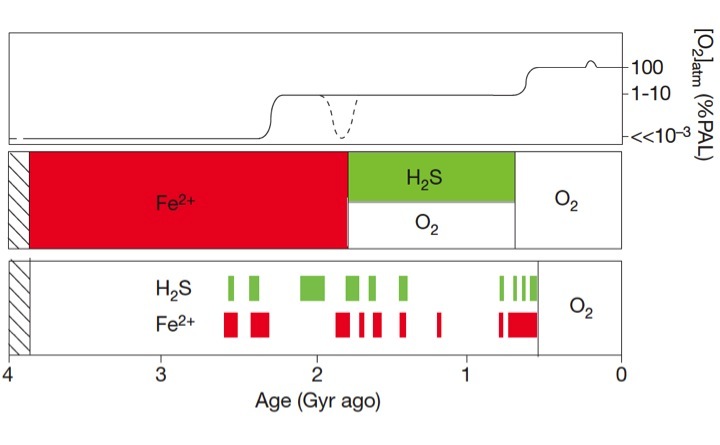
Summary of marine chemical conditions in the Precambrian ocean and atmosphere. a. Estimates of atmospheric oxygen concentrations relative to the present atmospheric level (PAL). b. Traditional models for the chemical composition of the deep ocean. c. Distribution of Precambrian euxinic and ferruginous deep waters based on our iron data from shales. Our data point to extensive and likely persistent ferruginous conditions in the deep ocean during the mid-Proterozoic, rather than the previously popular models favoring oxygenated or sulfidic conditions. The emerging view is that euxinic and ferruginous waters were simultaneously present during the mid-Proterozoic below oxygenated surface waters, with euxinia limited mostly to productive ocean margins. Spatial extents of euxinia were still likely great enough to impact the spatiotemporal distribution of prokaryotic and eukaryotic life by controlling nutrient availability. In the Phanerozoic, with higher levels of atmospheric oxygen, the deep oceans were anoxic only episodically and for short periods
Publications
-
Brennecka, G. A., Herrmann, A. D., Algeo, T. J., & Anbar, A. D. (2011). Rapid expansion of oceanic anoxia immediately before the end-Permian mass extinction. Proceedings of the National Academy of Sciences, 108(43), 17631–17634. doi:10.1073/pnas.1106039108
-
Dahl, T. W., Canfield, D. E., Rosing, M. T., Frei, R. E., Gordon, G. W., Knoll, A. H., & Anbar, A. D. (2011). Molybdenum evidence for expansive sulfidic water masses in ~750Ma oceans. Earth and Planetary Science Letters, 311(3-4), 264–274. doi:10.1016/j.epsl.2011.09.016
-
Dahl, T. W., Hammarlund, E. U., Anbar, A. D., Bond, D. P. G., Gill, B. C., Gordon, G. W., … Canfield, D. E. (2011). Reply to Butterfield: The Devonian radiation of large predatory fish coincided with elevated atmospheric oxygen levels. Proceedings of the National Academy of Sciences, 108(9), E29–E29. doi:10.1073/pnas.1018818108
-
Herrmann, A. D., Kendall, B., Algeo, T. J., Gordon, G. W., Wasylenki, L. E., & Anbar, A. D. (2012). Anomalous molybdenum isotope trends in Upper Pennsylvanian euxinic facies: Significance for use of δ98Mo as a global marine redox proxy. Chemical Geology, 324-325, 87–98. doi:10.1016/j.chemgeo.2012.05.013
-
Kendall, B., Anbar, A. D., Kappler, A., & Konhauser, K. O. (2012). The Global Iron Cycle. Fundamentals of Geobiology, None, 65–92. doi:10.1002/9781118280874.ch6
-
Konhauser, K. O., Lalonde, S. V., Planavsky, N. J., Pecoits, E., Lyons, T. W., Mojzsis, S. J., … Bekker, A. (2011). Aerobic bacterial pyrite oxidation and acid rock drainage during the Great Oxidation Event. Nature, 478(7369), 369–373. doi:10.1038/nature10511
-
Li, C., Love, G. D., Lyons, T. W., Scott, C. T., Feng, L., Huang, J., … Chu, X. (2012). Evidence for a redox stratified Cryogenian marine basin, Datangpo Formation, South China. Earth and Planetary Science Letters, 331-332, 246–256. doi:10.1016/j.epsl.2012.03.018
-
Liermann, L. J., Mathur, R., Wasylenki, L. E., Nuester, J., Anbar, A. D., & Brantley, S. L. (2011). Extent and isotopic composition of Fe and Mo release from two Pennsylvania shales in the presence of organic ligands and bacteria. Chemical Geology, 281(3-4), 167–180. doi:10.1016/j.chemgeo.2010.12.005
-
Loyd, S. J., Marenco, P. J., Hagadorn, J. W., Lyons, T. W., Kaufman, A. J., Sour-Tovar, F., & Corsetti, F. A. (2012). Sustained low marine sulfate concentrations from the Neoproterozoic to the Cambrian: Insights from carbonates of northwestern Mexico and eastern California. Earth and Planetary Science Letters, 339-340, 79–94. doi:10.1016/j.epsl.2012.05.032
-
Lyons, T. W. (2012). A perfect (geochemical) storm yielded exceptional fossils in the early ocean. Proceedings of the National Academy of Sciences, 109(14), 5138–5139. doi:10.1073/pnas.1202201109
-
Lyons, T. W., & Reinhard, C. T. (2011). Earth science: Sea change for the rise of oxygen. Nature, 478(7368), 194–195. doi:10.1038/478194a
-
Lyons, T. W., Reinhard, C. T., Love, G. D., & Xiao, S. (2012). Geobiology of the Proterozoic Eon. Fundamentals of Geobiology, None, 371–402. doi:10.1002/9781118280874.ch20
-
Mitchell, K., Mason, P. R. D., Van Cappellen, P., Johnson, T. M., Gill, B. C., Owens, J. D., … Lyons, T. W. (2012). Selenium as paleo-oceanographic proxy: A first assessment. Geochimica et Cosmochimica Acta, 89, 302–317. doi:10.1016/j.gca.2012.03.038
-
Owens, J. D., Lyons, T. W., Li, X., MacLeod, K. G., Gordon, G., Kuypers, M. M. M., … Severmann, S. (2012). Iron isotope and trace metal records of iron cycling in the proto-North Atlantic during the Cenomanian-Turonian oceanic anoxic event (OAE-2). Paleoceanography, 27(3), n/a–n/a. doi:10.1029/2012pa002328
-
Planavsky, N. J., McGoldrick, P., Scott, C. T., Li, C., Reinhard, C. T., Kelly, A. E., … Lyons, T. W. (2011). Widespread iron-rich conditions in the mid-Proterozoic ocean. Nature, 477(7365), 448–451. doi:10.1038/nature10327
-
Planavsky, N., Rouxel, O. J., Bekker, A., Hofmann, A., Little, C. T. S., & Lyons, T. W. (2012). Iron isotope composition of some Archean and Proterozoic iron formations. Geochimica et Cosmochimica Acta, 80, 158–169. doi:10.1016/j.gca.2011.12.001
-
Wasylenki, L. E., Weeks, C. L., Bargar, J. R., Spiro, T. G., Hein, J. R., & Anbar, A. D. (2011). The molecular mechanism of Mo isotope fractionation during adsorption to birnessite. Geochimica et Cosmochimica Acta, 75(17), 5019–5031. doi:10.1016/j.gca.2011.06.020
- Anbar, A.D. & Severmmann, S. (2011). Isotope Fractionation (Metal). In: Reitner, J. & Thiel, V. (Eds.). Encyclopedia of Geobiology. Springer.
- Anbar, A.D. & Severmmann, S. (2011). Transition Metals and their Isotopes. In: Gargaud, M. (Eds.). Encyclopedia of Astrobiology. Springer.
-
PROJECT INVESTIGATORS:
-
PROJECT MEMBERS:
Gordon Love
Co-Investigator
Chao Li
Collaborator
Amy Kelly
Postdoc
Brian Kendall
Postdoc
Kevin Nguyen
Doctoral Student
Noah Planavsky
Doctoral Student
-
RELATED OBJECTIVES:
Objective 4.1
Earth's early biosphere.
Objective 4.2
Production of complex life.

